1. 引言
如你所见,这是一类极不知名的生物,以至于整个中文互联网都几乎无法找到关于它的详细介绍(写完这篇之后算是有了)。即便是在学术界,它们也少有人关注。而在体型、种类、生存能力等各个方面,它们的表现都可谓中规中矩,正好印证了它们的名字——中阳虫($\text{Centrohelid}$)。

(上图:棘胞虫($Acanthocystis$),属中阳虫门针胞虫目,具有绵密的尖刺,广泛分布于淡水之中)
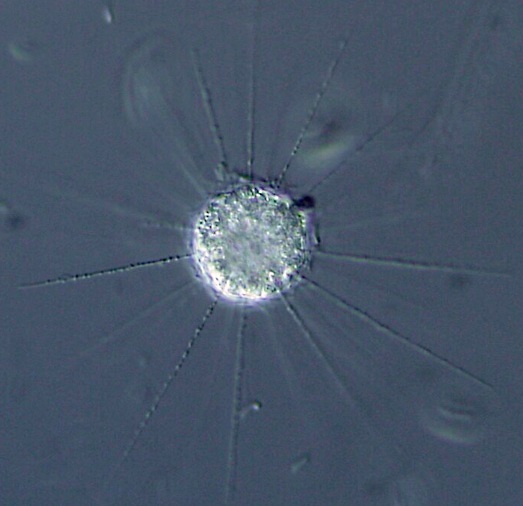
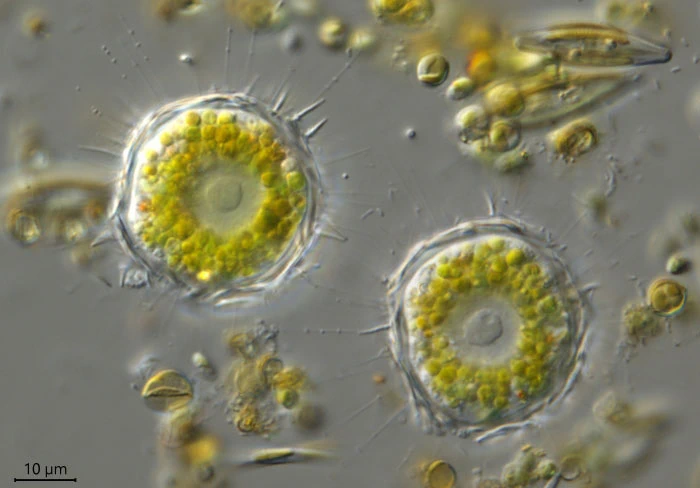
(上图:刺日虫($Raphidiophrys$),属中阳虫门刺日虫目,常见于淡水中)
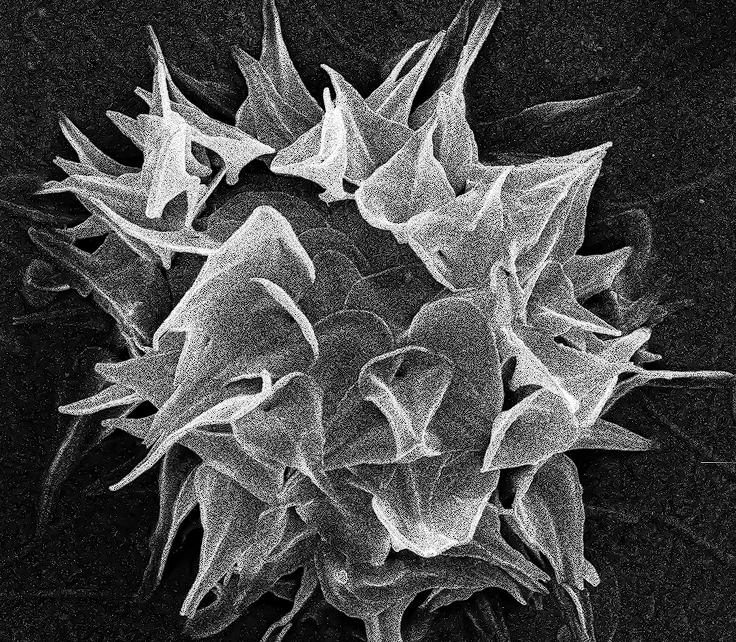
(上图:翼阳虫($Pterocystis$)的风干细胞照片,属中阳虫门翼阳虫目。其名称来源于特征性的翼状鳞片)
虫如其名,虽然个头只有几十微米大,但长得确实像一个小太阳。它们大多数淡水生物,体表披着一层硅质鳞片,并具有用于防御的尖刺。在很长一段时间里,它们都被归类到了一个名为太阳虫门($\text{Heliozoa}$)的分类群中。然而,在 $21$ 世纪分子生物学崛起之后,人们意识到太阳虫其实是一个多系群,其内部各类群都是独立演化成了这种形态,于是将其从正式分类群中删除。如今,中阳虫一般以独立的一门(中阳虫门,$\text{Centroheliozoa}$)被归入定鞭界($\text{Haptista}$)中。
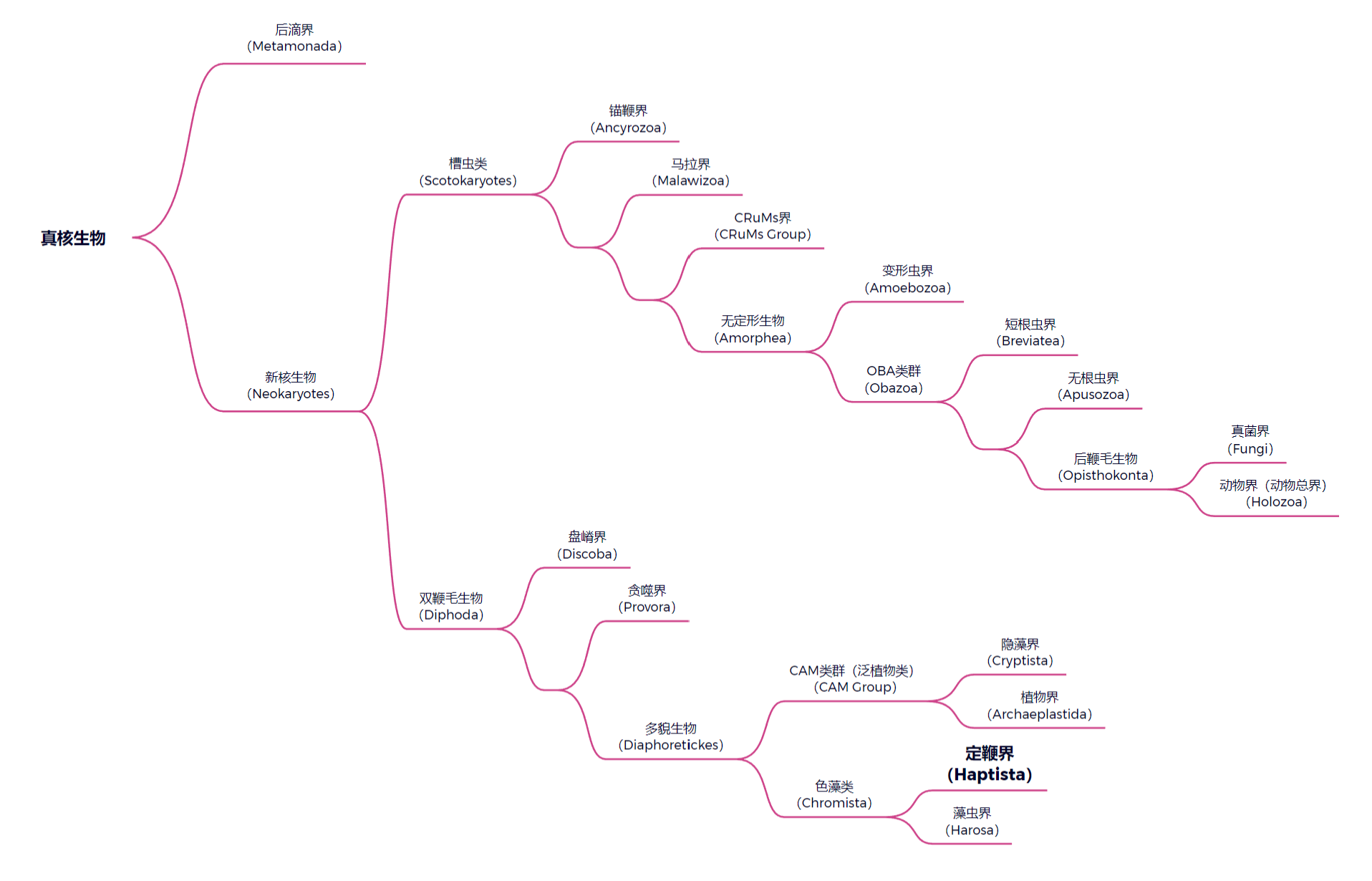
(上图:定鞭界的演化位置。需要注意,由于真核生物的系统发育树仍未完全确定,一些类群的位置会有争议。关于后滴界的问题,请见这篇文章)
分子生物学证据显示,中阳虫是定鞭界的基干分支,在 $\text{17-19}$ 亿年前即与其他生物分道扬镳。事实上,中阳虫可能是世界上第一类演化为太阳虫形的生物。这种形态不仅可以实现 $360^{\circ}$ 无死角防御,还对范围内所有的猎物都触手可及,真可谓无敌之身。可以说,太阳虫形是浮游伪足虫的最优解之一。但是,想要达到这种境界,也必然要付出代价。真核生物祖传的细胞骨架无法适配这种形态,因此各路太阳虫都对细胞结构进行了魔改,我们稍后也会讨论到。
然而,到了今天,这位元老似乎已经风光不再。如今最繁盛太阳虫的桂冠显然属于放射虫(放射虫门,$\text{Radiozoa}$),它们不仅可以长到零点几个毫米(注:它是真正的单细胞生物),分布范围还遍布全球海洋,是浮游生物的重要组成部分。它们的化石从寒武纪到现在,$5$ 亿多年来就没断过,将它们的繁盛展露无遗。

(上图:各种各样的放射虫)
反观中阳虫,不仅体型大小只有几十微米级,而且在它的各种生境中都有强力的竞争对手。诸如核形虫(核形虫门,$\text{Nuclearida}$)、粒缕虫(粒缕虫纲,$\text{Granofilosea}$)、针胞藻(针胞藻纲,$\text{Raphidophyceae}$)等选手的实力均不容小觑。虽然这些妥妥的都是太阳虫中的晚辈,但年轻气盛的它们一直在与中阳虫这位老同志争抢。
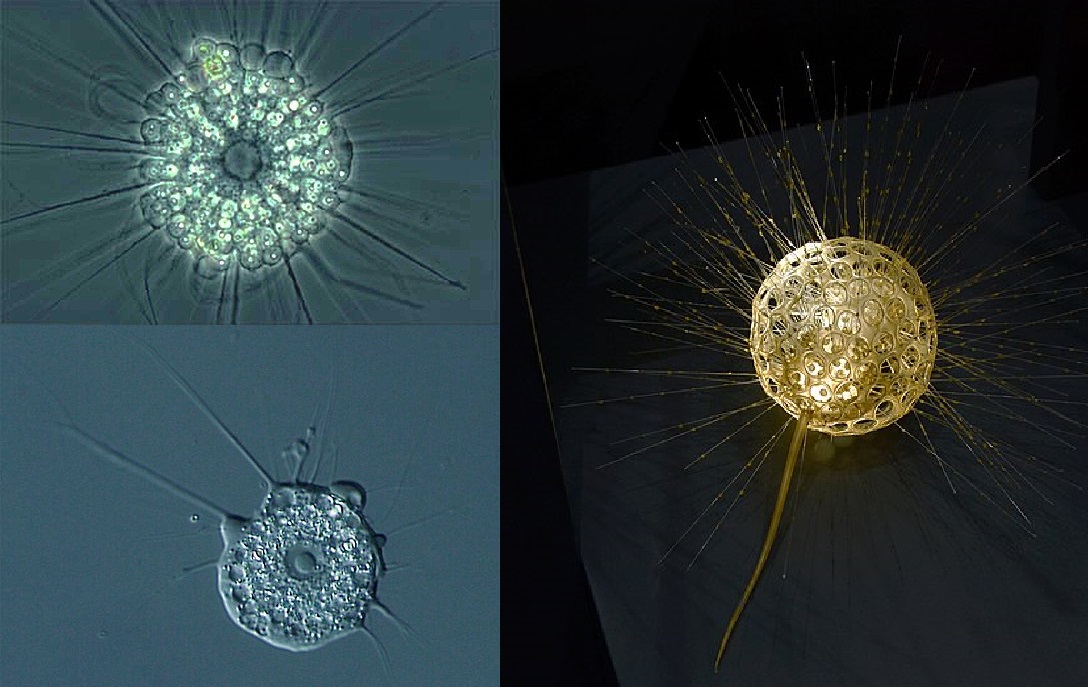
(上图左上:核阳虫目($\text{Actinophryida}$),属针胞藻纲;左下:核形虫;右:阳缕虫目($\text{Desmothoracida}$),属粒缕虫纲)
但反过来想,中阳虫混得也不算太差,毕竟也有十几科二十属的多样性,且很可能有许多物种我们还未发现。至少相比微阳虫(微阳虫门,$\text{Microheliozoa}$)来说是这样,这个门只有一个已知物种,体表裸露,体型只有几微米大,且于 $2012$ 年才被发现。分子证据显示,微阳虫是隐藻界($\text{Cryptista}$)的基干分支,分化时间也极早,甚至可以与中阳虫竞争一下谁才是太阳虫元老。它的身体结构也与中阳虫非常相似,二者可能是趋同演化。但不管怎样,它的处境比中阳虫悲惨得多。
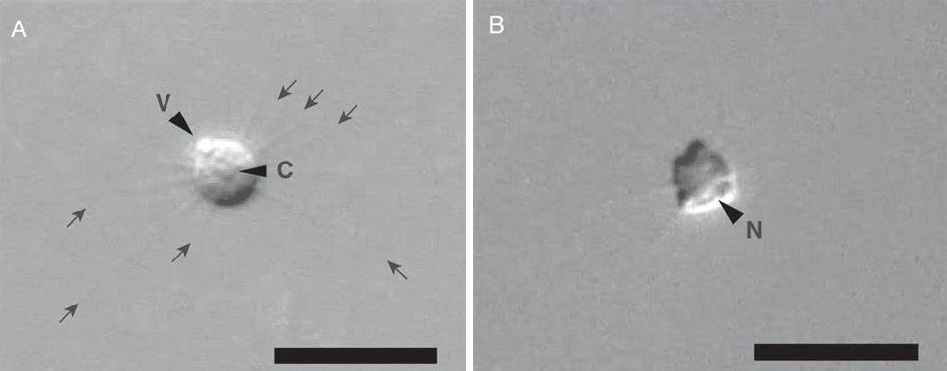
(上图:已知唯一一种微阳虫 $M.~maris$)
所以,今天的它们为什么会呈现出这样一种比上不足,比下有余的姿态呢?难道在十几亿年的悠长岁月中,中阳虫一直都固步自封,无所作为吗?并非如此。事实上,它们的生境横跨海陆,生存策略灵活多变,甚至还有一些支系将分散的个体团结起来,并开始向多细胞生物的方向演变。在下文中,我们将一起去看看这支默默无闻的族裔背后的生存之道。
2. 正文
前文提到,为了适配自己的形态,各路太阳虫都对自己的细胞结构进行了魔改,但即使是在这群魔乱舞中,中阳虫也属于改得最厉害的一派。我们先来回想一下,一个常规真核细胞的中心是什么?当然是细胞核,毕竟这可是遗传物质的存储地,指挥着整个细胞的生命活动;细胞核外则包裹着内质网,负责蛋白质合成等重要任务;此外还有高尔基体、中心体等等一堆细胞器。这些结构,应该都是天经地义的吧?
带着这种认知,让我们来看一眼中阳虫的结构。
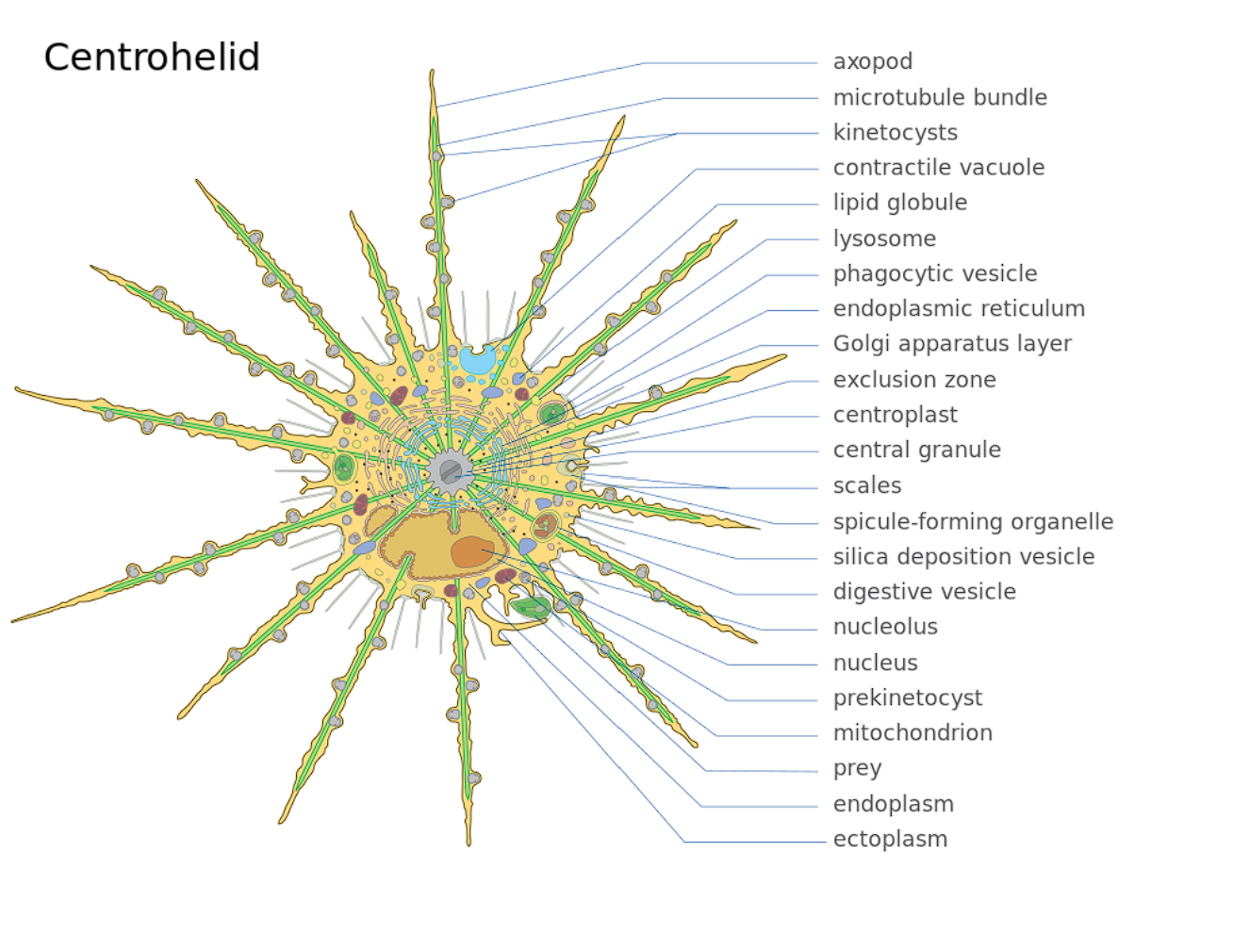
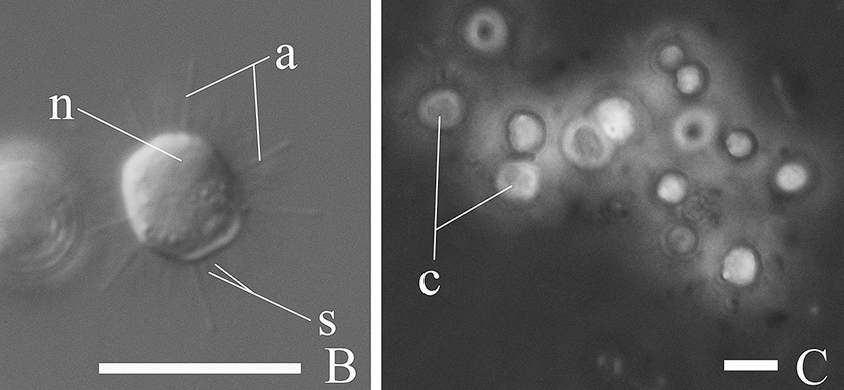
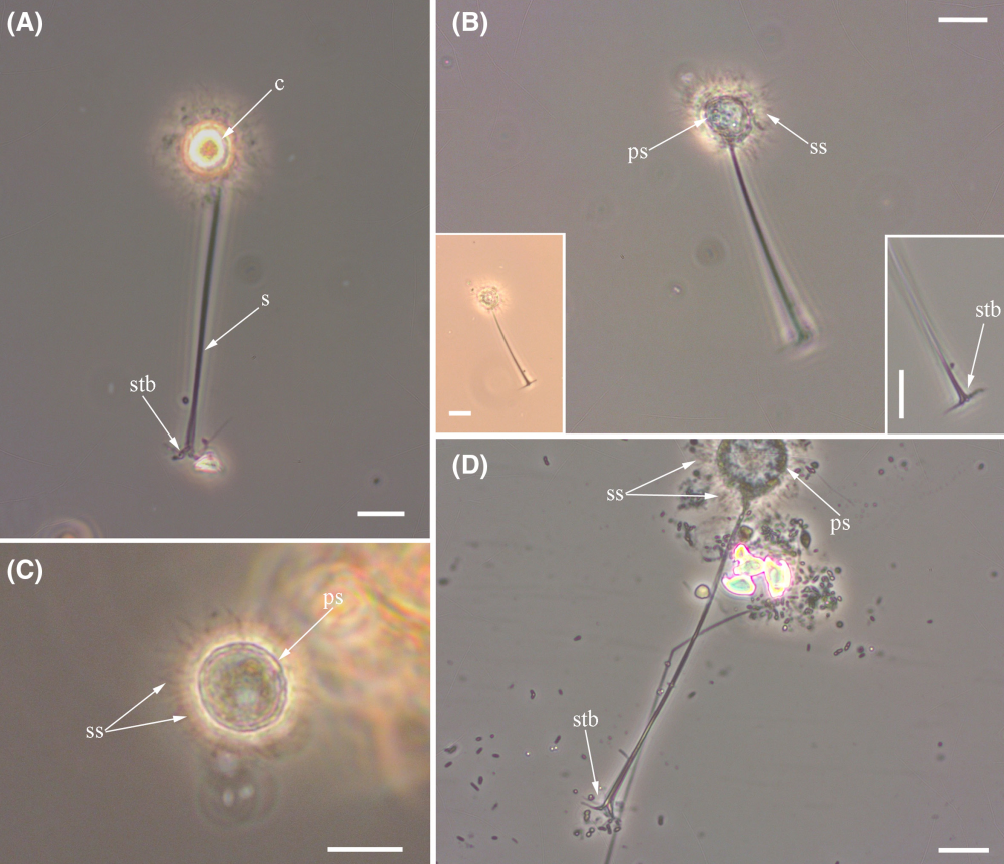
首先映入眼帘的当然是它的轴伪足($\text{axopod}$,简称轴足),这是太阳虫的经典结构,主要用于收集食物;其内部有用于支撑的微管阵列,即微管束($\text{microtubule bundle}$)。听起来很高大上,但其实就是把一堆微管捆成特定的形状罢了。仔细一看,轴足上还有一个个小点,像是悬挂在线上的珍珠。这便是动囊体($\text{kinetocysts}$),一种中阳虫特有的分泌型细胞器,具有精巧而复杂的结构。当轴足捕捉到猎物时,动囊体便将其内部的毒素注入猎物体内,使它无法反抗。
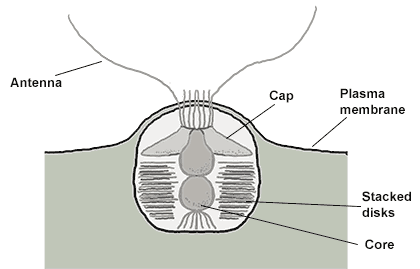
(上图:动囊体的结构简笔。整体呈圆形或锥形,两条须($\text{Antenna}$)朝向猎物的位置)
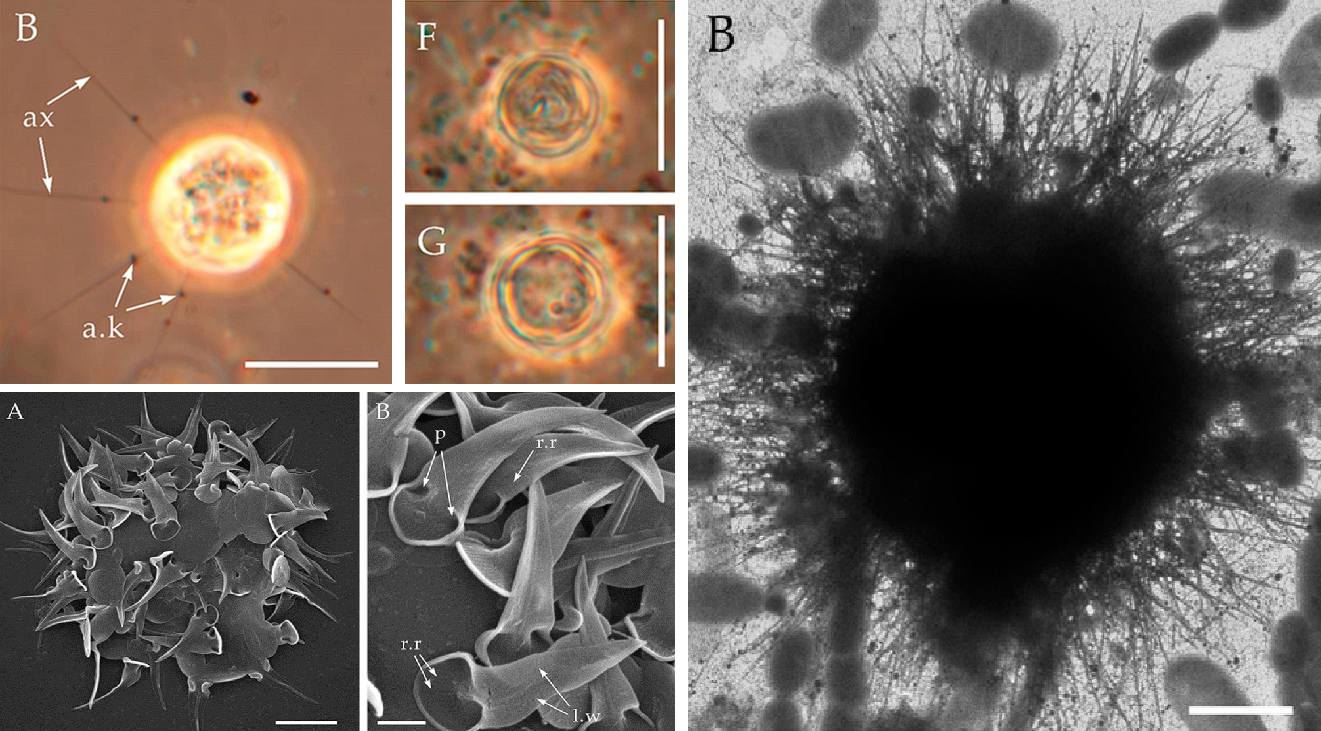
到此为止,一切似乎还挺正常,但当我们把视线移向中央时,情况却不对劲了起来。中阳虫的细胞中心并非细胞核,而是一个被称为中心质($\text{centroplast}$)的结构;而中心质的中心,是一个被称为中心颗粒($\text{central granule}$)的球体。早在 $19$ 世纪中叶,也就是中阳虫刚刚被发现那会,人们就已经注意到了这些特殊结构。后来又有人注意到,中心质深度参与了中阳虫的有丝分裂,且它的复制过程与动物细胞的中心体($\text{centrosome}$)如出一辙。所以一般认为,中心质和中心体是同源的细胞器。
遗憾的是,我们今天仍无法表述清楚这些神奇结构的具体功能,但它们显然与微管束系统密切相关,且可能充当了整个系统的支点,承受了来自四面八方的压力;我们也相信,演化既然把这些结构放到了细胞中心,那它们一定在中阳虫的生命活动中有着非同一般的地位。
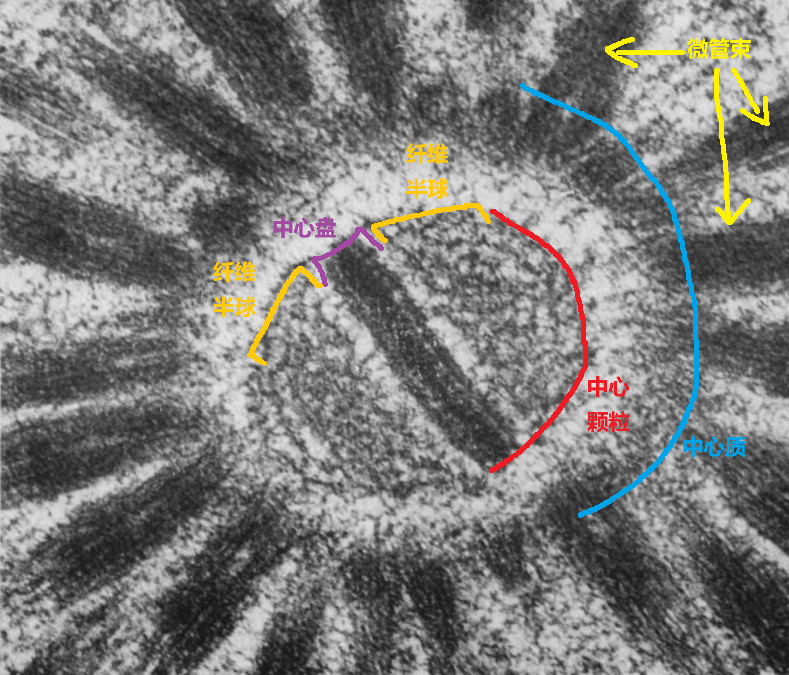
(上图:中阳虫 $H.~elati$ 细胞中心结构的显微图片。可以看到,中心颗粒被分为三叶,最中间的称为中心盘($\text{central disk}$),结构致密;另外两叶是由纤维状物质构成的半球)
在中心质之外,有一层没有任何细胞器的奇妙空间,被称为“禁区”($\text{exclusion zone}$),功能未知(或许是阻止外界物质干扰中心质?);而在“禁区”之外,包裹着高尔基体层,再外面才是内质网。没错,中阳虫把高尔基体团成了一个空心球,并将其包到了内质网里面(可能是为了配合中心质工作?);而细胞核只能委屈地缩在内质网的一侧。
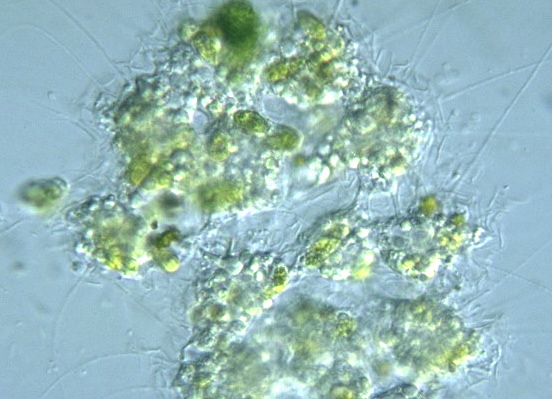
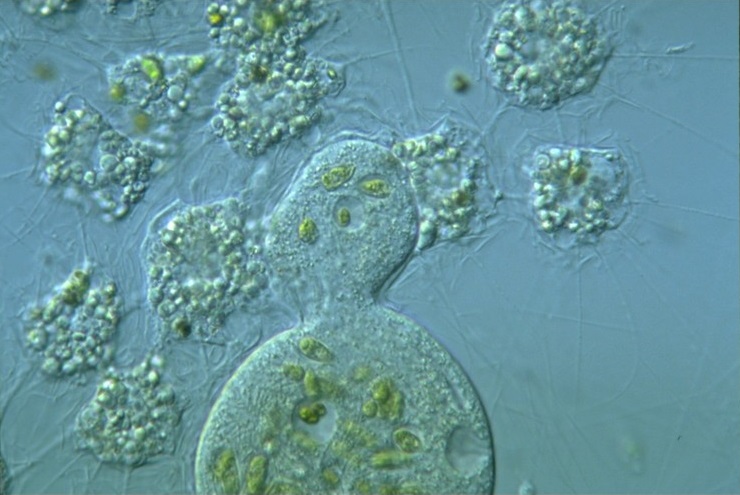
总的来说,中阳虫的细胞结构是高度特化的,而这幅身躯也让它们成为了那个微小世界中顶级的掠食者。当然,它们有时也会圈养一些藻类来进行光合作用,也算是经典操作了。
(上图:中阳虫捕食 $C.~elongatum$(一种绿藻)的珍贵影像(但估计是投喂的))
(上图:内共生了小球藻($Chlorella$,绿藻门)的 $A.~turfacea$)
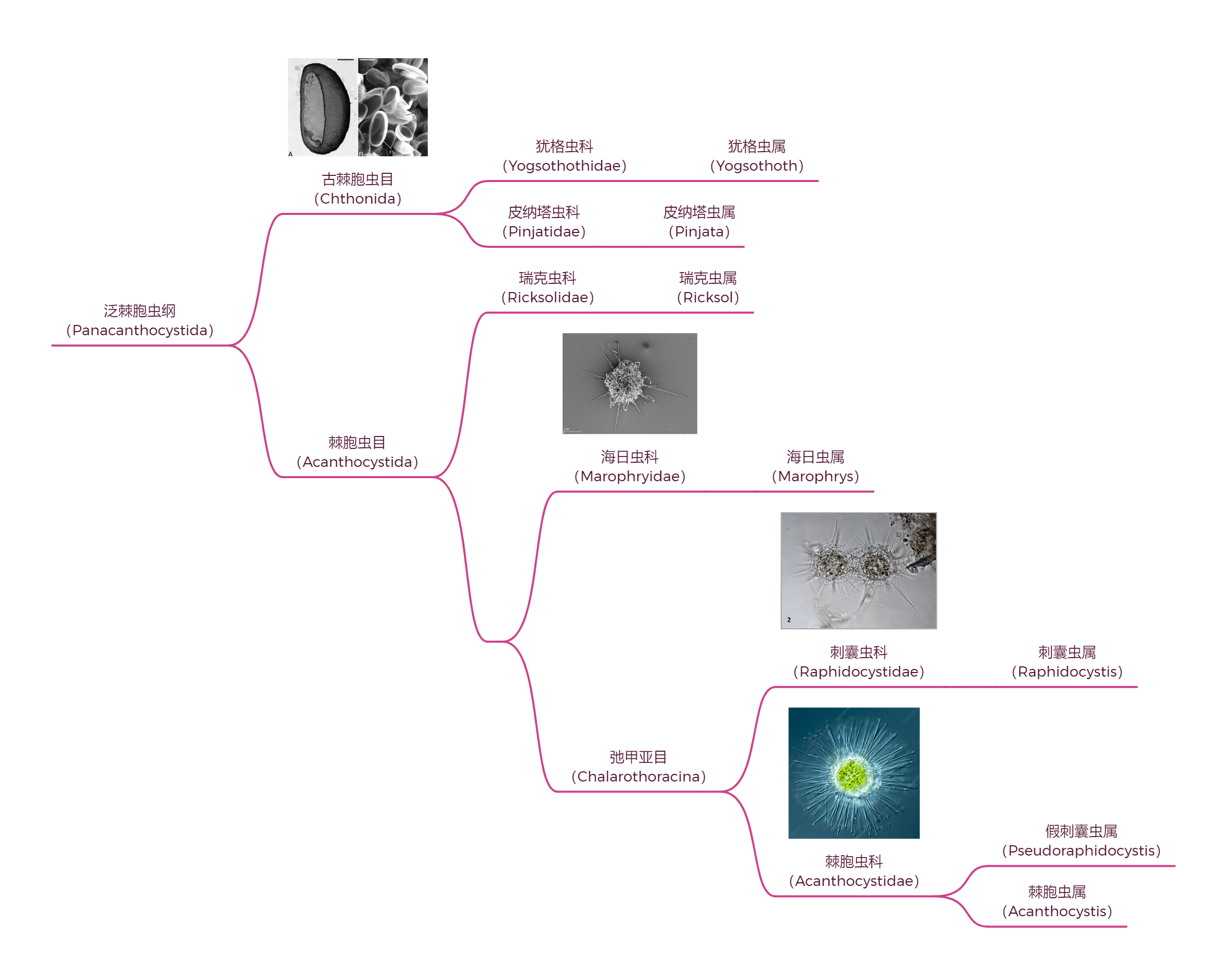
中阳虫的内部分类一直以来比较混乱,即使是 $21$ 世纪分子生物学出手之后,也依然存在很大争议。此处仅展示一种(我个人最相信的)观点,即门内部可分为两纲五目,如下图所示:
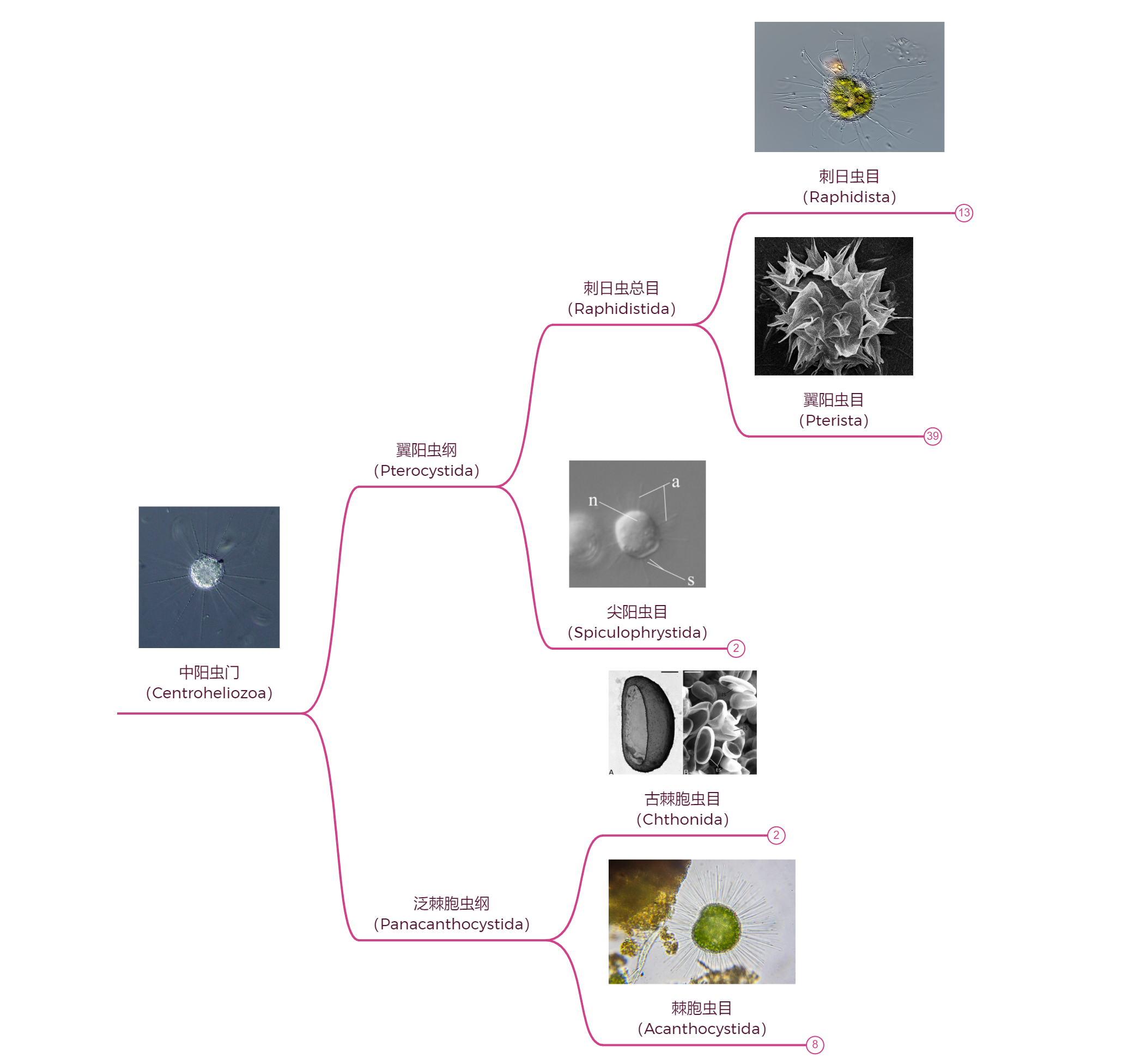
翼阳虫纲($\text{Pterocystida}$)是两纲中较大的一个。位于纲基部的尖阳虫目($\text{Spiculophrystida}$)只有一个属,即 $2016$ 年被发现的尖阳虫属($Spiculophrys$)。与很多中阳虫不同,尖阳虫的体表没有硅质鳞片,但有一种锐利的尖刺,由有机质构成。在此之前,我们曾认为硅质鳞片在整个中阳虫门的最近共祖那儿就已经演化成型了,而这一认知已经被尖阳虫的发现打破。目前的主流观点认为,中阳虫在早期演化中发生了两次独立的硅化事件,而尖阳虫的形态可能很接近中阳虫的祖先。
尖阳虫的另一个不同之处是,它生活在海洋之中,但不是表层浮游生物,而是栖息在 $100m$ 以下的海域中,这或许也是中阳虫祖先的栖息地。在细胞培养中,经常可以观察到它们聚在一起形成群落,这应该是它们独立演化出来的生存策略:
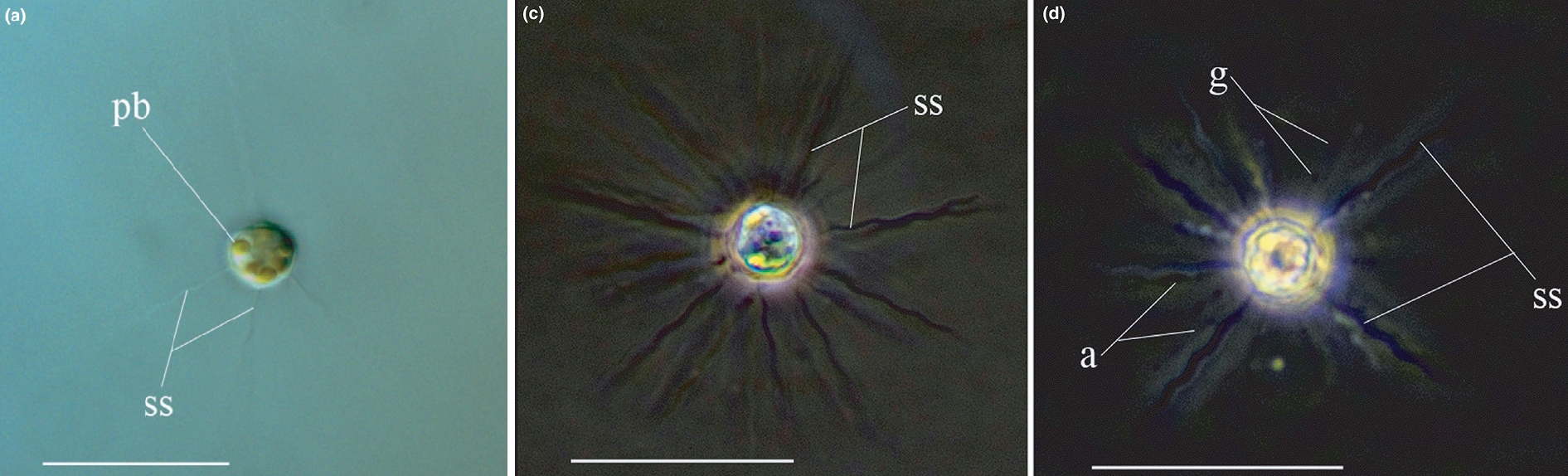
(上图:尖阳虫属的 $S.~agregata$ 的个体细胞(左)及群落(右))
翼阳虫纲的另一分支是刺日虫总目($\text{Raphidistida}$),中阳虫的其中一次硅化可能就发生在该类群的祖先那里。其下两目中,刺日虫目($\text{Raphidista}$)相对更知名一些,如下图所示:
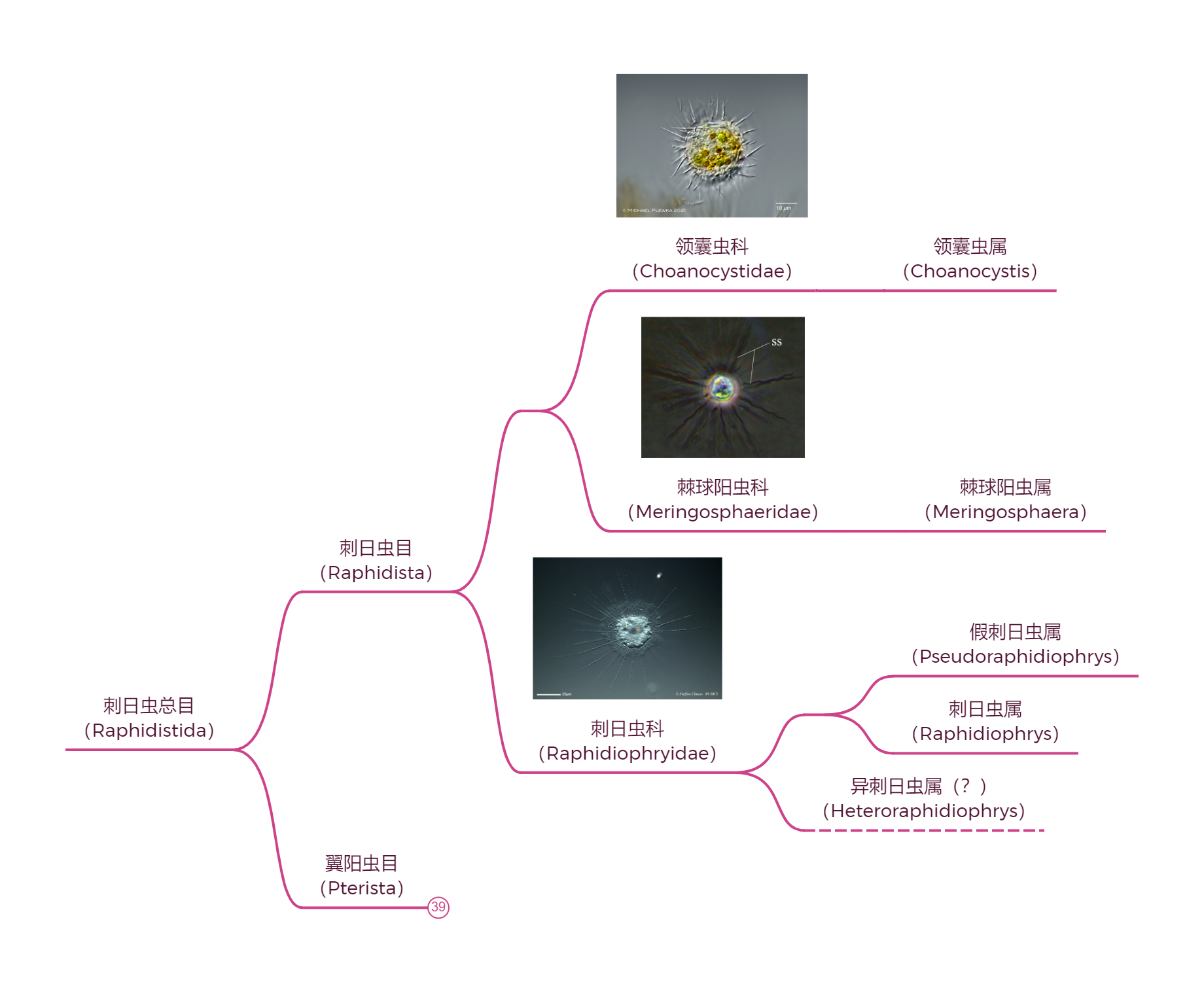
刺日虫($Raphidiophrys$)或许是少数几种你可能听闻过的中阳虫。它们广泛分布在世界各地的淡水中,具有多样的生存策略。个体既可以单干,也可以像尖阳虫那样形成群落(一般仅有几个细胞,但也有罕见的毫米级甚至接近一厘米的大群落),从而常态化地捕食草履虫等较大的猎物。它们的群落中可以观察到胞质桥,用于个体之间的物质交换。有些时候,它们也会与藻类共生来进行光合作用,自给自足。
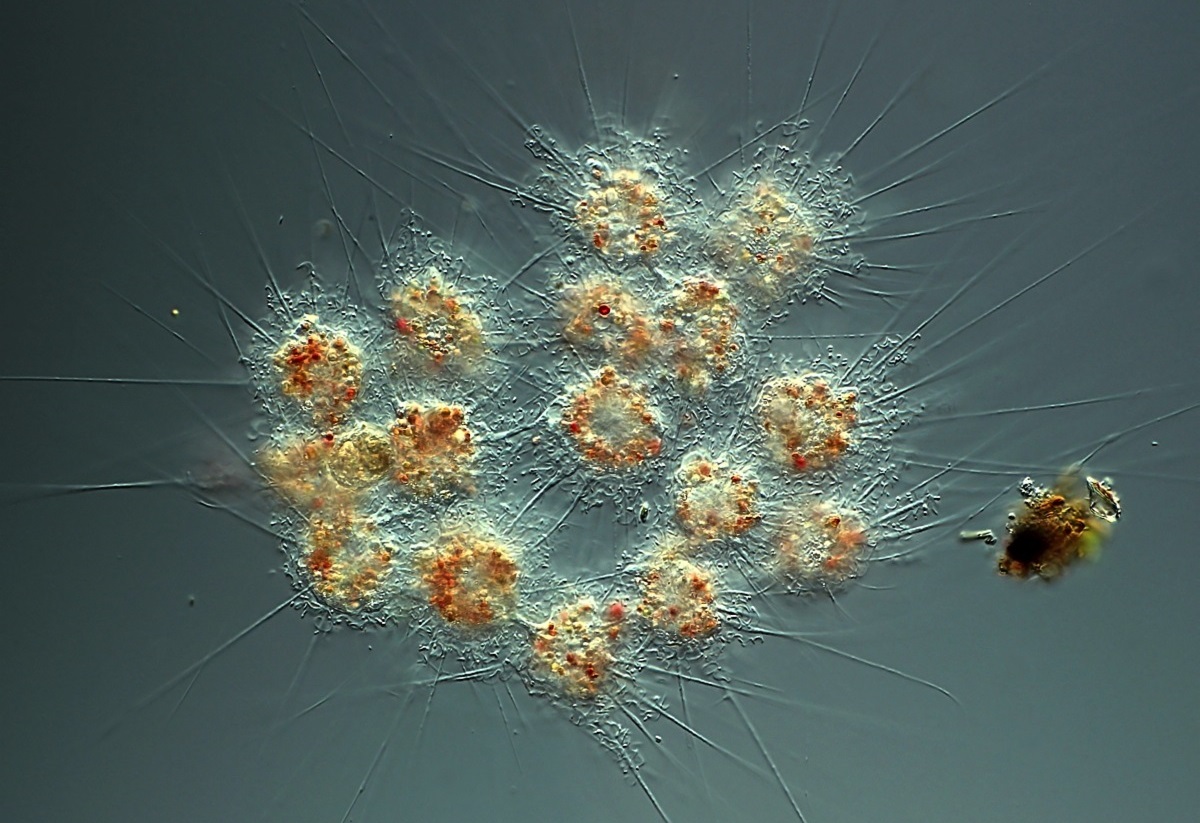
(上图:$R.~elegans$ 形成的一个不算大的群落,颜色来自于共生的藻类)
它们的硅质鳞片像个鞋垫一样扁平,仔细一看,鳞片由上下两块板构成(这被认为是该属物种的标志),中间是空心的,由一块块隔板($septa$)支撑;隔板弯曲的部分向外突出,形成所谓的肋拱($rib$)结构。这样的结构不仅轻薄,而且强度很高,同时也比实心的结构更加节省硅质。
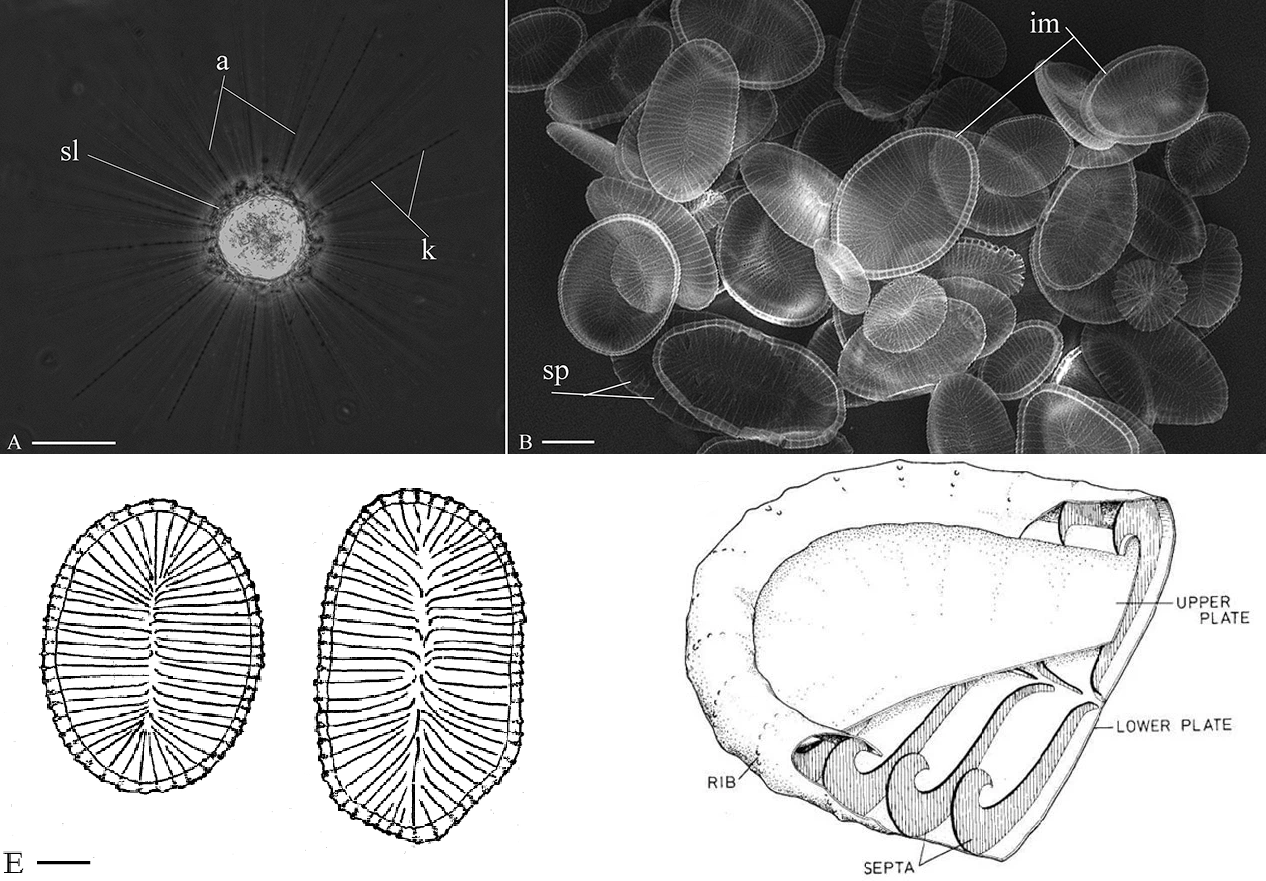
(上图左上:$R.~drakena$ 单个细胞活体;右上:$R.~drakena$ 的鳞片;左下:鳞片外观手绘;右下:鳞片内部结构手绘)
与刺日虫关系很近的假刺日虫($Pseudoraphidiophrys$)虽然个体体型更小,但在群体协作上毫不逊色。它们几乎不会独自行动,只要周围有同伴就会自发聚集为群落。它们的群落也更加多样,除了刺日虫同款的松散联盟(仅通过轴足、黏液和胞质桥纠缠在一起)外,还有一个将每个细胞都紧密地贴在一起的特殊形态(接近我们通常认知上的“多细胞”形态)。但是,这很可能只是一种过渡状态,因为假刺日虫还发展出了合胞体。
所谓合胞体,即是一种大型的多核细胞,由一堆细胞融合而成。假刺日虫的群落和合胞体的规模一般不大,在 $10$ 个细胞左右,但也有一些多达上百个细胞,尺寸达到毫米级:
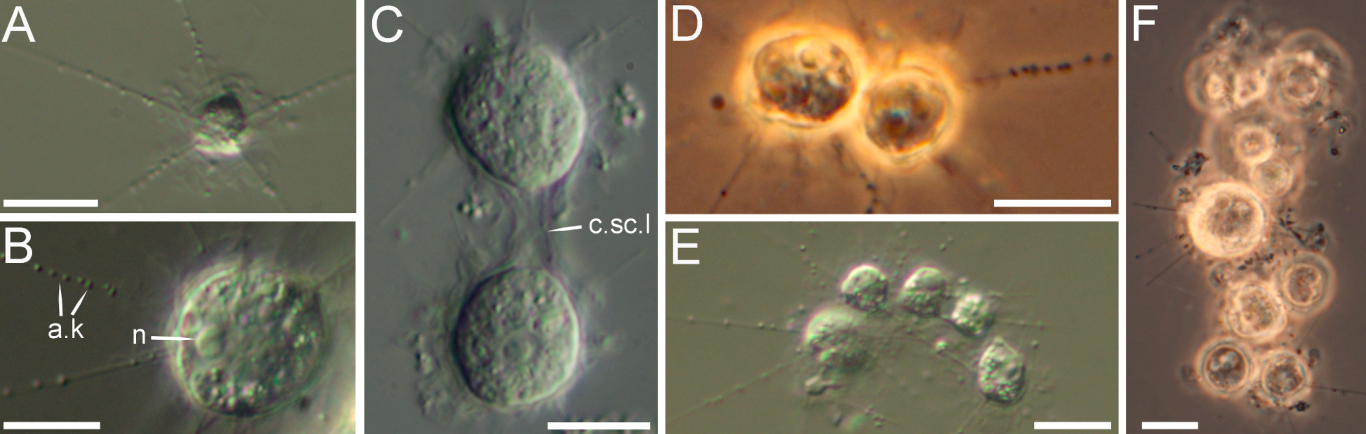
(上图:$P.~veliformis$。$\text{A,B}$:单个细胞;$\text{C}$:正在分裂的细胞;$\text{D,E}$:正在聚集的细胞;$\text{F}$:已初具规模的群落)
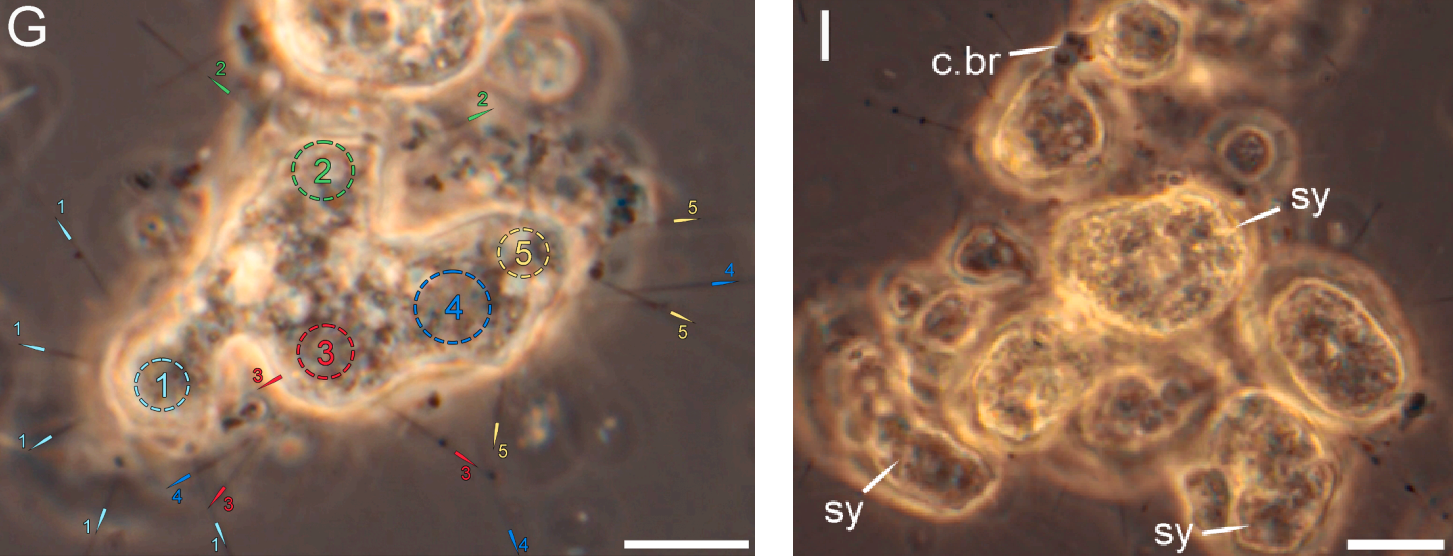
(上图 $\text{G}$:一个合胞体的部分,可以观察到 $5$ 个融合为一体的细胞;$\text{I}$:一个特殊的群落,十多个细胞紧密地贴在一起)
在一些群落里,还能观察到一种带柄的假刺日虫个体。这种结构的作用尚不可知,但有一种猜想:利用这种结构,群落中的一些个体可以把轴足伸向更广阔的水域,从而捕获更多猎物;柄起到了类似安全索的作用,使得该个体不会脱离群落。当条件恶劣时,假刺日虫的群落会解体,散作满天星斗;群落中的每个细胞都会形成包囊,进入休眠状态。包囊没有轴足,但具有加厚的细胞膜和包裹全身的宽厚鳞片(而且一般会包上好几层),以保护细胞本体。
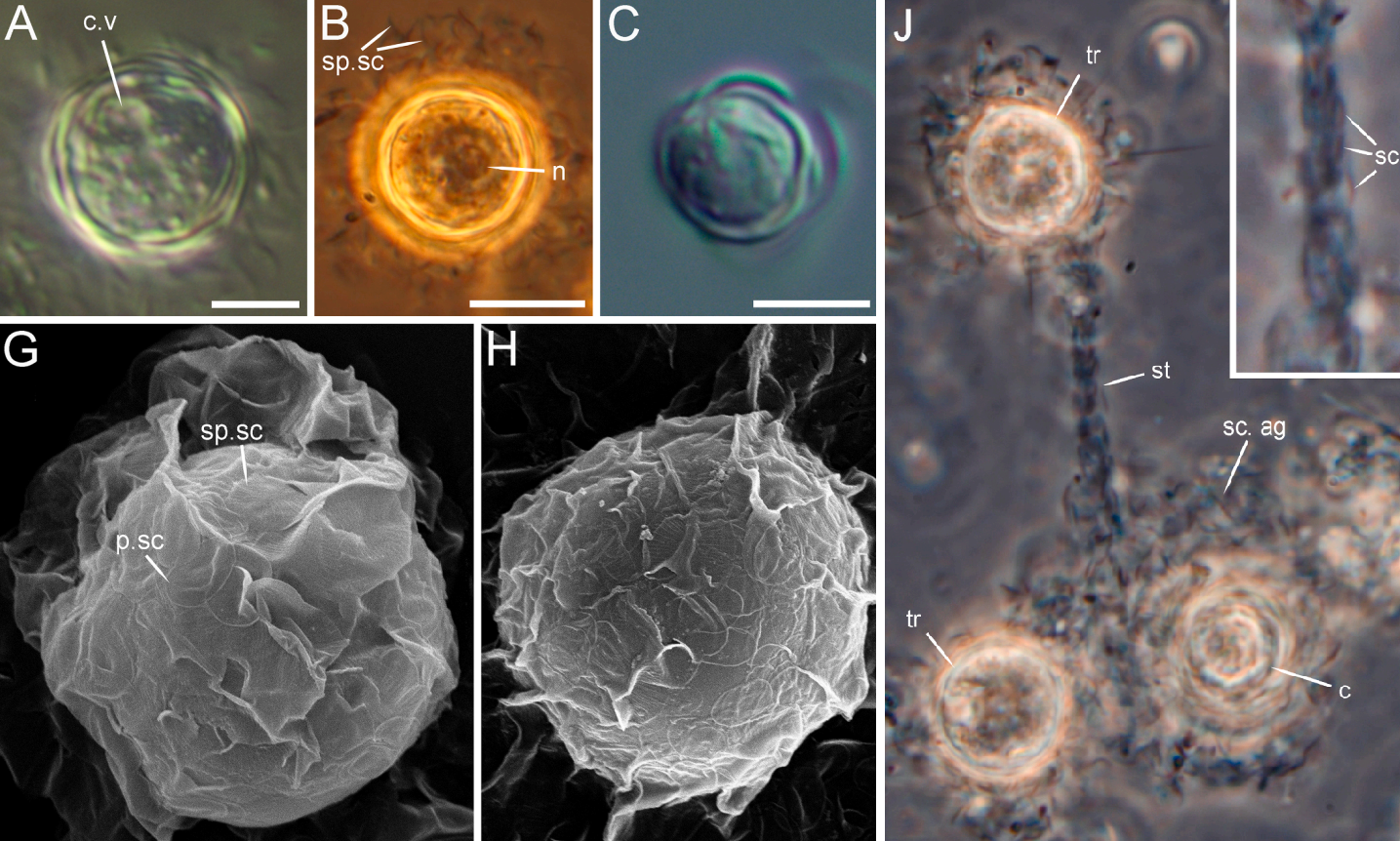
(上图 $\text{A-C}$:包囊;$\text{G-H}$:包囊表面微结构的特写;$\text{J}$:一个小群落,其中有带柄的个体)
刺日虫和假刺日虫两属合在一起,可以构成刺日虫科($\text{Raphidiophryidae}$),是刺日虫目的基干分支;目下还有另外两个类群,其中的棘球阳虫($Meringosphaera$)在很长一段时间里都被认为是一类金藻,直到 $2021$ 年的一项研究才将其确认为中阳虫。在显微镜下,它们的细胞确实泛着金光;它们生活在浅海中,且所有个体无一例外都有内共生的藻类:
(上图:$M.~mediterranea$)
领囊虫($Choanocystis$)是棘球阳虫的姐妹,其下有几十个物种,世界性分布。它们早在 $19$ 世纪就被发现,体表有绵密的刺,在淡水和海洋中均可以找到。有意思的是,该属内成员的体型差异很大,既有可以长到 $70μm$ 的 $C.~pantopoda$,也有细胞直径仅 $3μm$ 的已知最小的中阳虫 $C.~minima$:
(上图:$C.~minima$,风干细胞照片)
(上图:$C.~aculeata$,最常见的领囊虫,生活于淡水中)
刺日虫总目的另一类群——翼阳虫目($\text{Pterista}$)则是门内最大的目级演化支,如下图所示:
甲阳虫($Clypifer$)于 $2021$ 年被发现,是翼阳虫目的基群。它们生活在近海中,体型也比较小,平均细胞直径为 $6μm$。它们的鳞片是刺日虫同款的鞋垫状,但没有后者那样精巧的超微结构;中央有一个明显的凹陷区域,还有一根短轴连接两翼,又使它有了点翼阳虫的味道:

(上图:$C.~cribrifer$)
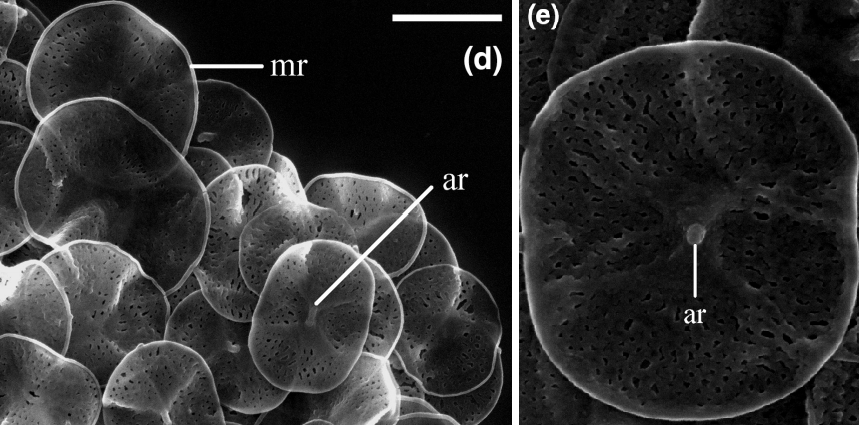
(上图:$C.~cribrifer$ 的鳞片)
相比于甲阳虫,奥阳虫($Oxnerella$)在树上的位置更深,但也更加奇特。没错,它们既没有刺,也没有硅质鳞片,甚至连粘连鳞片的黏液都退化掉了,成为了中阳虫中唯一的裸太阳虫(这是要和微阳虫看齐了?)。它们同样也生活在海洋中,目前只有两个物种被发现,且体型较小,只能靠吃细菌为生:
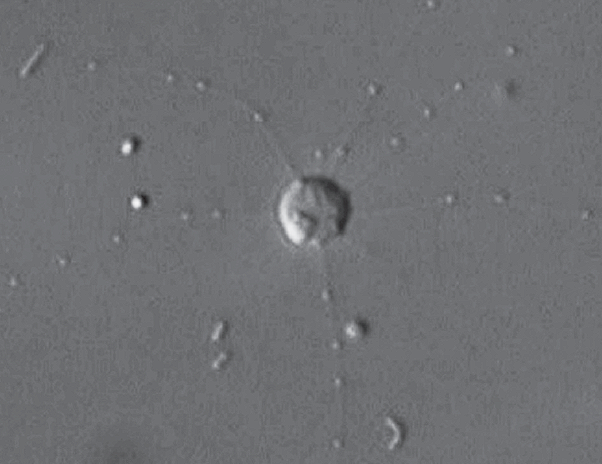
(上图:$O.~maritima$,可见其细胞表面明显地与其他中阳虫不同)
翼阳虫目中绝大多数物种都在真翼阳虫总科($\text{Eupterocystidae}$),但其内部的分类真可谓一言难尽。这个问题还得怪翼阳虫属($Pterocystis$),这个拥有三十多个物种的大属是一个超级并系群(甚至可能是多系群):
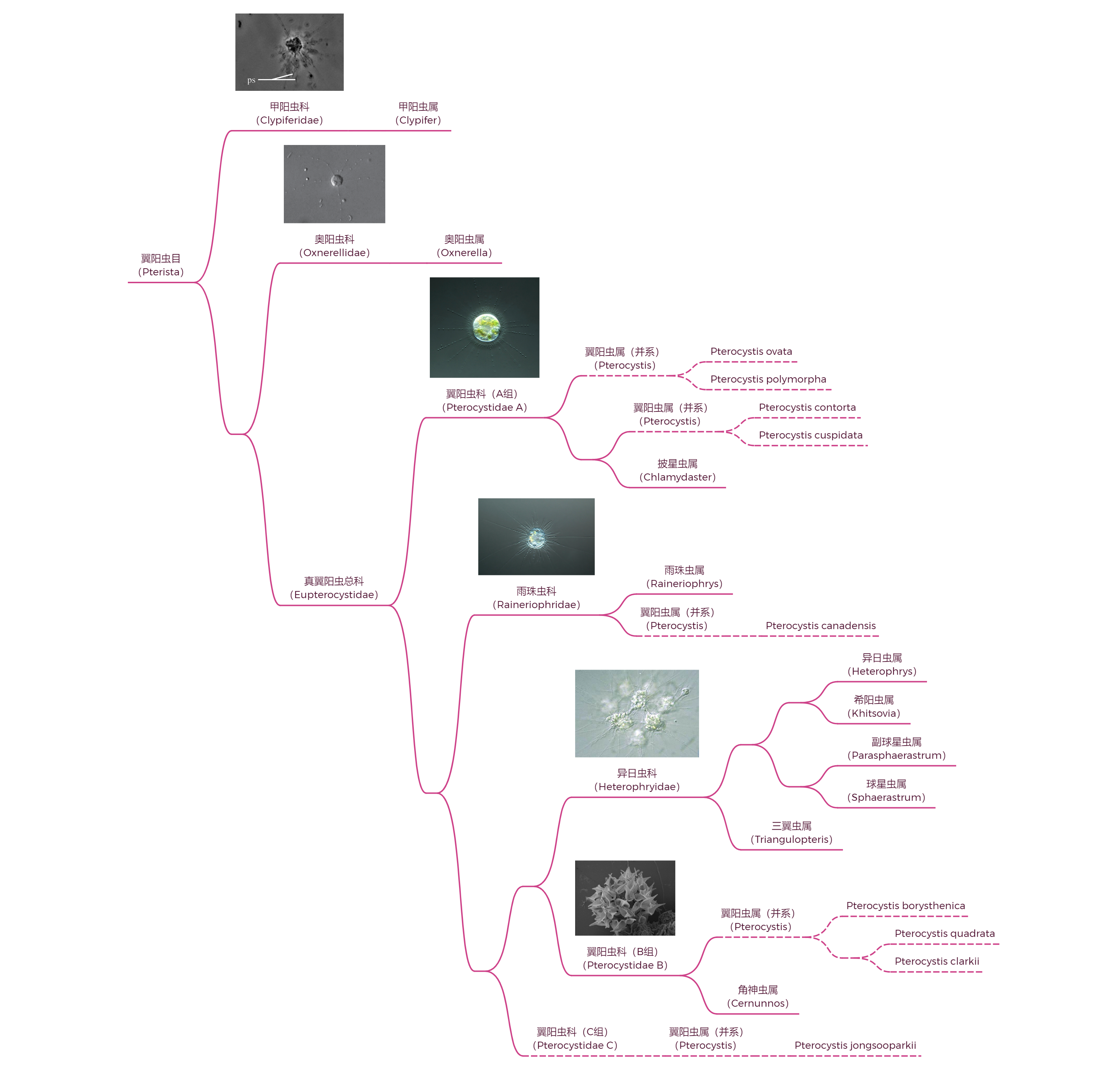
该属需要至少分成五组(一些研究中更多),才能维持其单系性。在形态学上,这是非常匪夷所思的,因为翼阳虫属具有明显的特征,即翼状的鳞片。
(上图:$P.~polycristalepis$。$\text{A,B}$:活细胞;$\text{C}$:风干细胞;$\text{D}$:鳞片特写)
翼阳虫的翼状鳞片中间凹陷,两边上翘,顶端尖锐,可以充当尖刺扎伤来犯之敌。但是,伸展的两翼大大增加了鳞片占用的空间,使其无法单独铺满细胞表面。所以,翼阳虫还拥有第二种形态的鳞片,一般是扁盘状,用以确保细胞表面没有漏风之处。可以说,这样的鳞片设计是极为精巧的;也正是因此,人们理所当然地认为翼阳虫是单系群。而分子生物学的发现便是告诉我们,自然界总是没有我们所想得那么简单。
目前,关于翼阳虫属何去何从的问题,学术界也没有定论,所以我们也不再纠结这个问题。
在翼阳虫科($\text{Pterocystidae}$,自然也不是单系群)中,还有一个属唤作披星虫($Chlamydaster$)。这货没有鳞片,也没有尖刺,但它并不是裸太阳虫。与奥阳虫不同,披星虫用黏液(原本是用于粘住鳞片的)把自己包了起来,形成了被称为黏液套的结构,并在表面布设了一堆绒毛,就像是给自己织了一层毛衣:

(上图:$C.~laciniatus$ 的简笔画(左)与活细胞(右))
我们至今仍不知道披星虫为何会这样演化,更难以理解其黏液和绒毛的具体作用,因为相比于尖刺,这些结构就像是在给敌人挠痒痒。
雨珠虫($Raineriophrys$)的形态则更为奇特。它的细胞外面有很多弯弯曲曲缠绕在一起的细长而柔软的“面条”(好吧我实在想不出用什么词可以描述这种结构),且即使形成包囊,这些“面条”依然会被保留。一般状态下,它的轴足非常长,可以达到细胞直径的五倍以上:
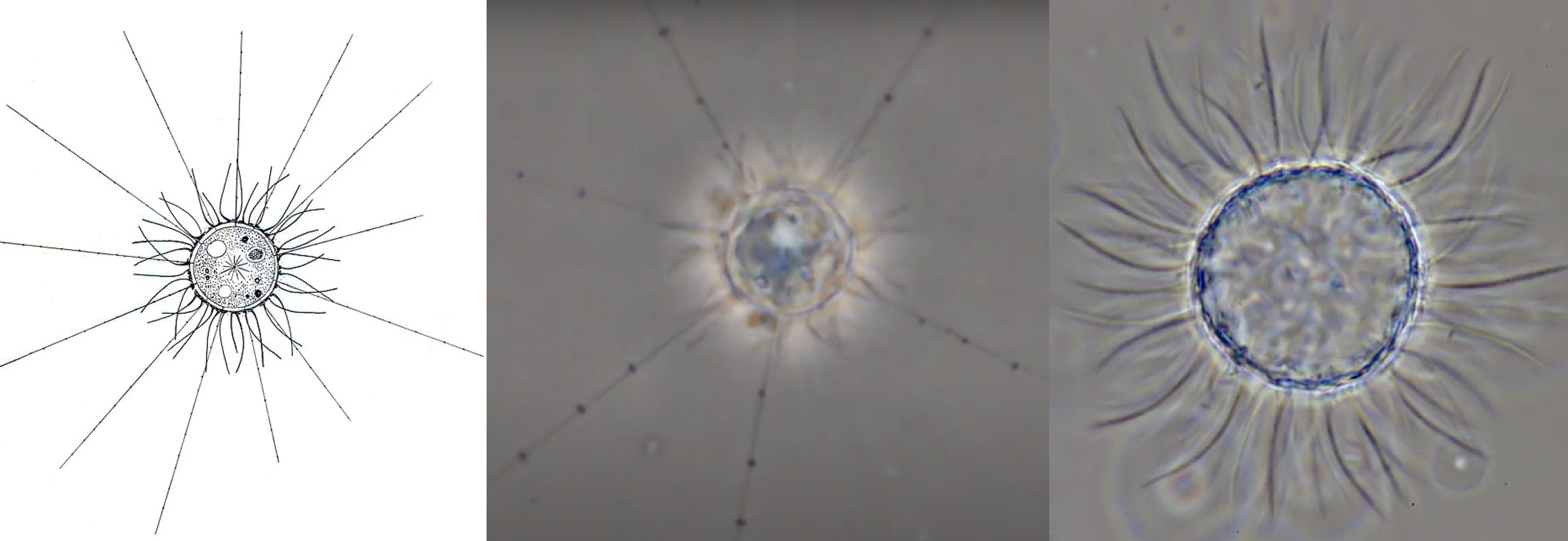
(上图:$R.~echinata$ 的简笔画(左)、正常活细胞(中)与包囊(右))
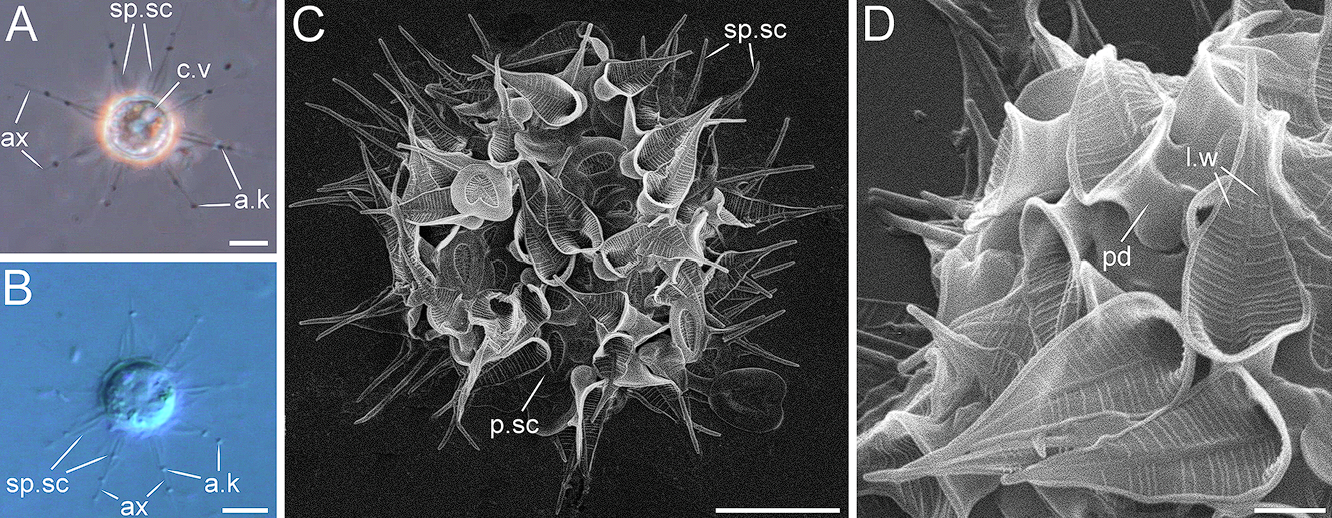
角神虫($Cernunnos$)则是 $2023$ 年新发现的类群。它有一种非常特殊的尖锐鳞片,具有一个明显的弯曲主干和许多细小的分叉,看起来像鹿角一般,也因此被冠以凯尔特神话中长着鹿角的神 $\text{Cernunnos}$ 之名。以此为基础,$C.~arctica$ 还演化出了一根长长的柄,从而得以固着在基质之上:
(上图:$C.~arctica$;图 $\text{C}$ 为俯视,所以看不到柄)
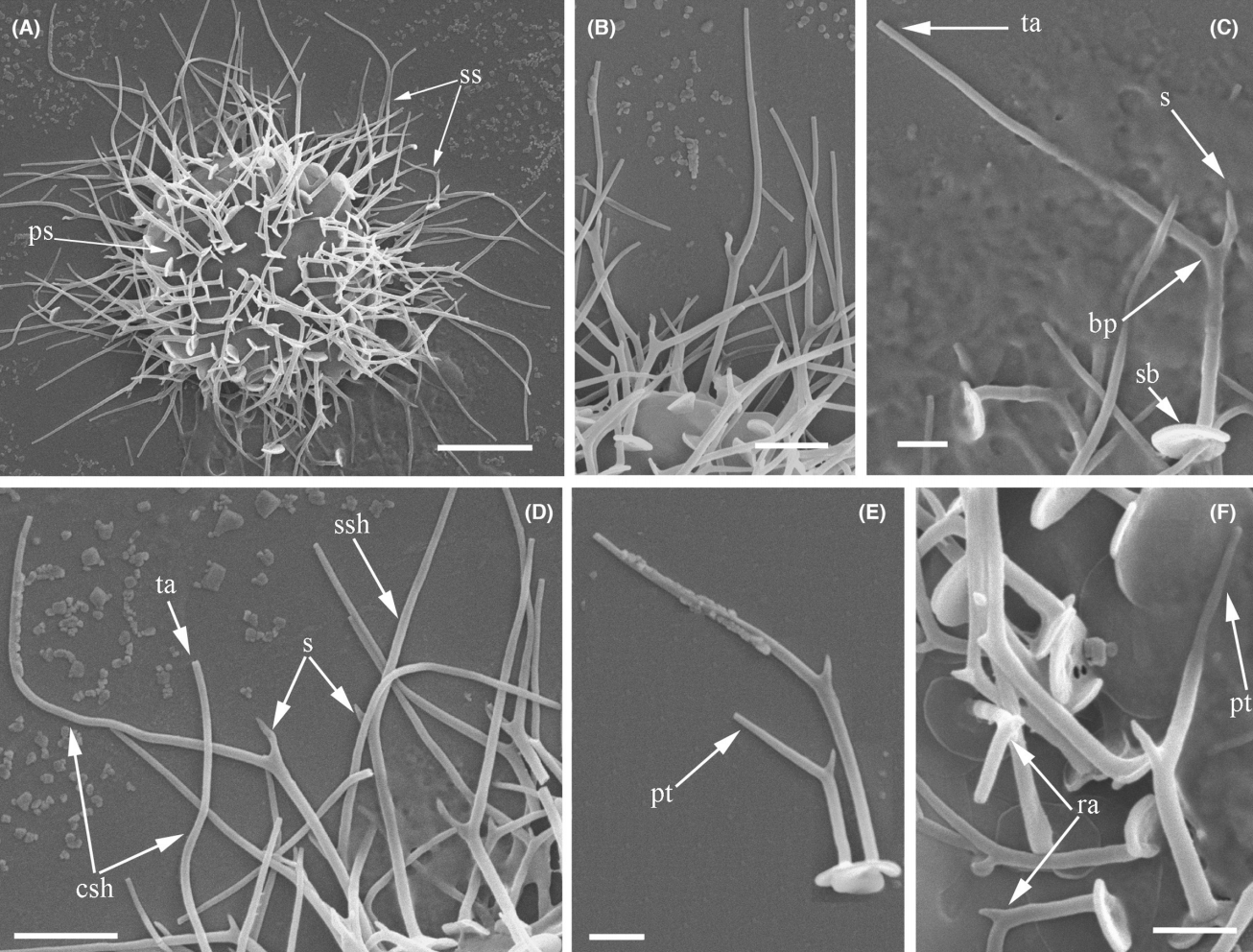
(上图:$C.~uralica$ 的风干细胞($\text{A}$)与尖锐鳞片特写($\text{B-F}$))
此处插一嘴,本文中所说的“鳞片”无论被改造成什么样子,是尖的也好平的也罢,主要成分都是硅质;而所谓“尖刺”,是由某些有机物构成的,这也是区分二者的方法。
从下属类群数量来看的话,异日虫科($\text{Heterophryidae}$)才是翼阳虫目中最大的类群。位于科基部的三翼虫($Triangulopteris$)被发现于土壤中,它们的鳞片是弯曲的三角形,尖锐的顶端与翼阳虫有异曲同工之妙。此外,在培养中还观察到它们有一种浑身长刺但没有鳞片的形态,乍一看宛如返祖成尖阳虫了一般,但这正是异日虫科的特征:
(上图:$T.~lacunata$,左上 $\text{B}$:活细胞;$\text{F,G}$:包囊;左下 $\text{A}$:风干细胞;中下 $\text{B}$:鳞片特写;右侧 $\text{B}$:尖刺形态的风干细胞)
处于演化树更深位置的球星虫($Sphaerastrum$)生活于淡水,已经完全褪去了鳞片,转而采用黏液套加尖刺的防御手段。它们又一次独立演化出了群落,规模也可达毫米级,并学会了用黏液套和胞质桥连接每个个体:
(上图:$S.~fockei$)
(上图:正在与另一只原生生物战斗的 $S.~fockei$ 群落)
副球星虫($Parasphaerastrum$)则堪称整个中阳虫门中最神秘的类群,自从 $1996$ 年被首次发现以后便再也没有被记录过,以至于我连张活细胞照片都没找到。它被认为是球星虫的姐妹,生活在海洋中。其表面尖刺很有特色,是切向的,就像一堆针贴在细胞表面:
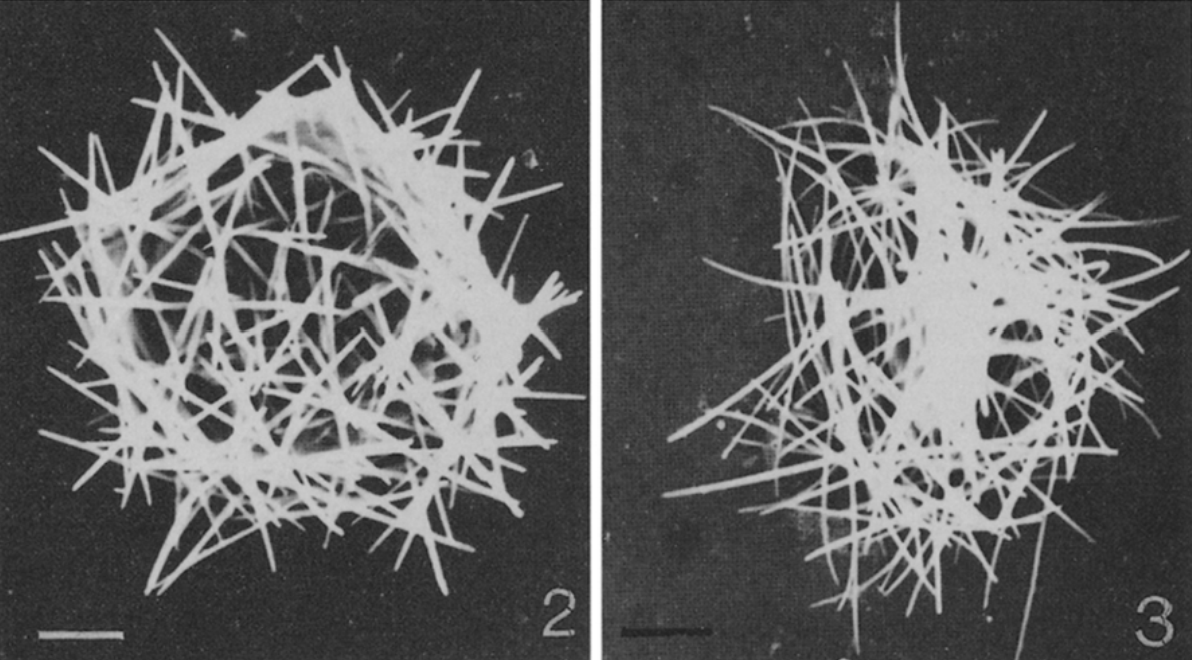
(上图:$P.~marina$ 风干细胞(左)与散落的尖刺(右))
异日虫科的大当家——异日虫属($Heterophrys$)一样是黏液套配尖刺,但它不会形成群落,且所有尖刺都是法向的,看起来锋芒毕露。它生活在淡水中,体型也比较大,可以长到 $70μm$ 以上:
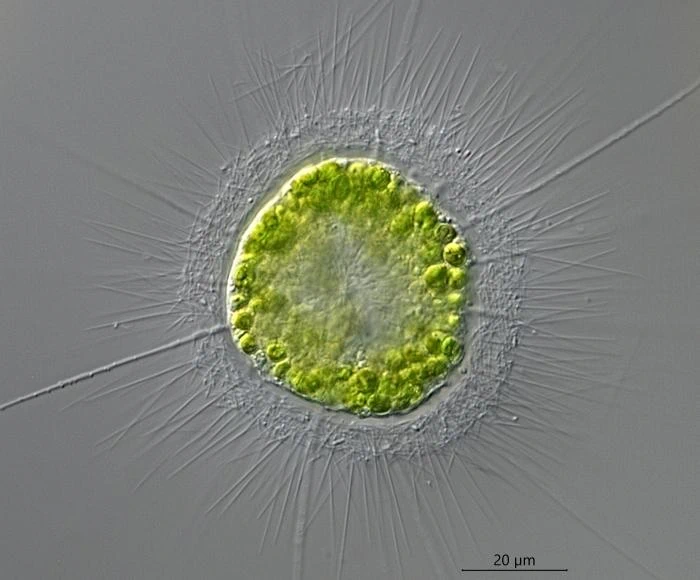
(上图:$H.~myriopoda$,最长的那几条是轴足,短而密的是尖刺)
异日虫属的姐妹希阳虫($Khitsovia$)于 $2022$ 年被发现于滩涂和泥沼中。粗看起来,它反而与三翼虫比较相似,具有鳞片和尖刺双形态。有趣的是,其部分鳞片像是一个勺子,且“勺柄”部分是根空心管,顶端还留有开口;另有许多扁盘状的鳞片,以确保细胞被完全覆盖:
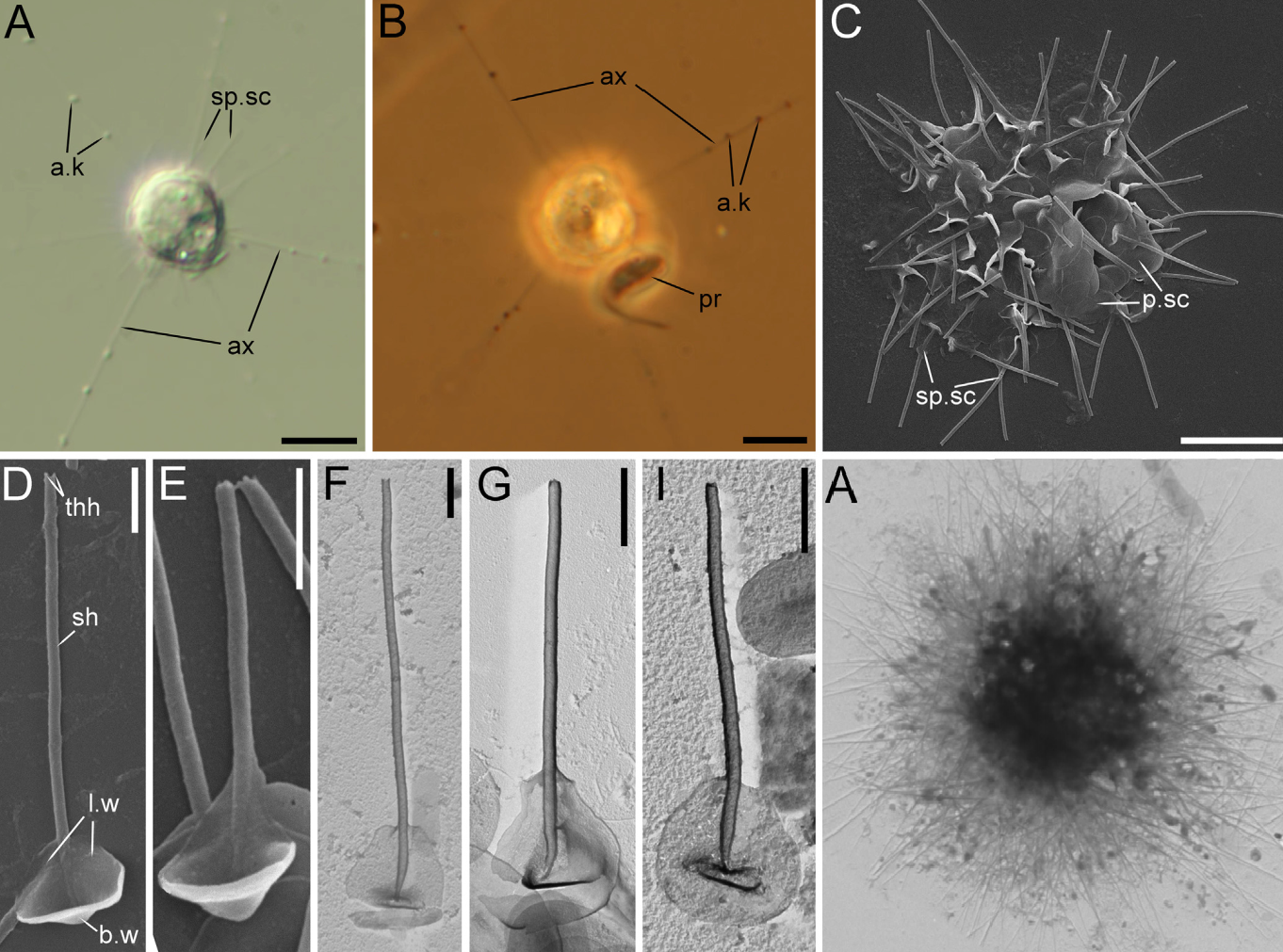
(上图:$K.~mutabilis$,$\text{A,B}$:活细胞;$\text{C}$:风干细胞;$\text{D-I}$:勺状鳞片特写;右下 $\text{A}$:尖刺形态的风干细胞)
至此,我们算是把中阳虫门中最大的一纲看了个大概,而另一纲——泛棘胞虫纲($\text{Panacanthocystida}$)相对来说要小得多,其下仅有 $7$ 属。但是,这并不代表它们就是演化的漏网之鱼,相反,可能是因为在正常的道路上没什么竞争力,它们中的很多都点出了一堆匪夷所思的黑科技,令人大开眼界。
棘胞虫属($Acanthocystis$)是纲内最早被发现的类群,也是纲名的来源。很多地方把它翻译为“刺胞虫”,但我觉得这个译名无论从哪个角度来看都很差劲。从词源看,属名的前半部分来自拉丁语 $\text{acanthus}$,意为“荆棘”;从实际运用来看,“刺胞”这一名字明显与刺胞动物($\text{Cnidaria}$)撞车。因此,虽然这一译名广为流传,但在本文中不会使用。
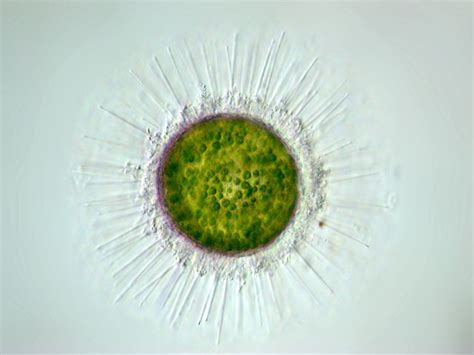
言归正传,棘胞虫可能是整个中阳虫门中最繁盛的属,不仅有近四十个物种,而且生境横跨海陆,在野外的湖泊、河流中都能轻易找到。这首先要归功于它们的护甲。棘胞虫的细胞外不仅有密密麻麻的法向尖刺(可以秒杀密集恐惧症的那种),还有两种不同的鳞片:一种扁平,覆盖细胞表面;另一种尖锐,顶端带有分叉,但长度比尖刺更短。这还不够,它们在鳞片下又用蛋白质铺设了一层坚韧的保护膜,被称为周质体。如此四合一的防御体系,真可谓登峰造极:
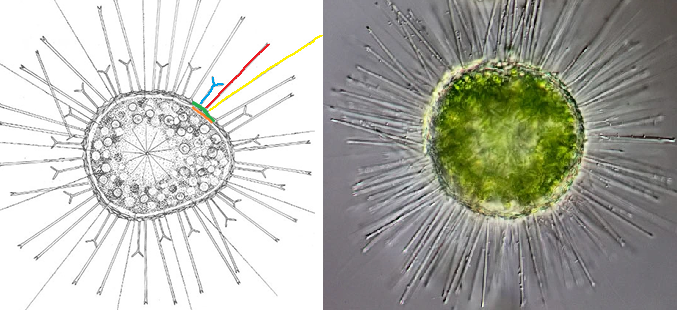
(上图:$A.~turfacea$;左侧简笔画中,黄色为轴足,红色为尖刺,蓝色为特化的尖锐鳞片,绿色为扁平鳞片层(部分),橙色为周质体(部分))
另外,它们对内共生藻类的掌控也更为得心应手,基本属于随心所欲,想光合作用就光合作用,不想光合作用就直接杀了吃的水平。它们内共生的藻类也更加多样,使得它们在显微镜下可以呈现出绿色、黄色、黄绿色甚至是褐色,当然也有无色的:
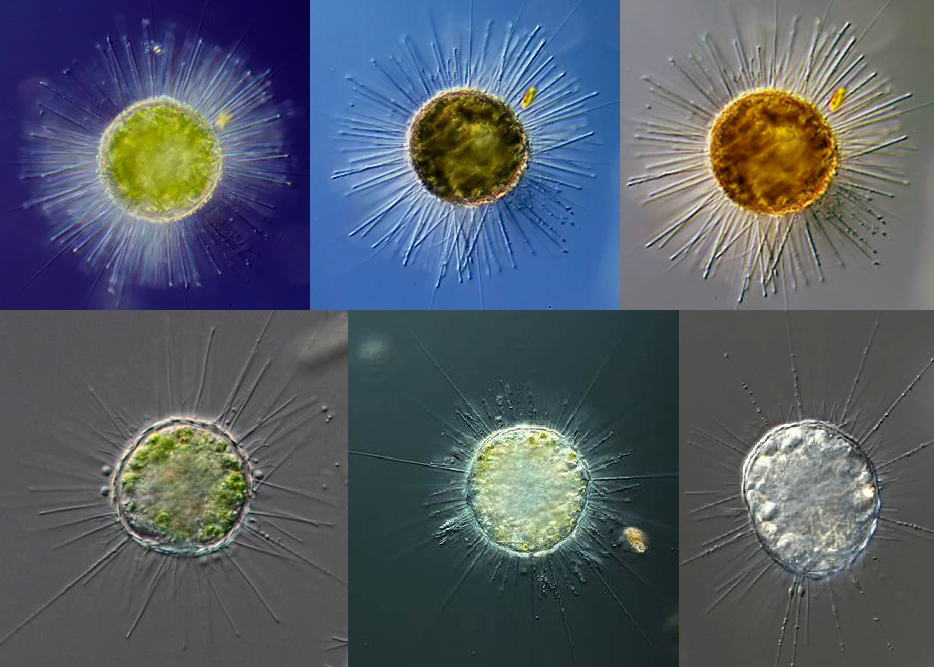
(上图:共生了不同藻类的棘胞虫;最后一张图中没有明显的内共生藻类)
同样较早被发现的是刺囊虫($Raphidocystis$)。它的名字和刺日虫很像,形态上更像,若是不看鳞片的超微结构则几乎无法靠肉眼区分,但它们其实根本不是一路的,亲缘关系也极远,二者不过是趋同演化罢了。事实上,有许多刺囊虫都曾被错误地归为刺日虫,直到近几年才改过来(具体见第三章节)。
作为全方位的山寨货,刺囊虫不仅学习了刺日虫的外观,还学习了它们的集团化策略。不少刺囊虫都可以形成群落,同时保持着单独生活的能力。另一些如 $R.~tubifera$ 借鉴了棘胞虫的设计,也把一部分鳞片变得尖锐,还在顶端添上了分叉:
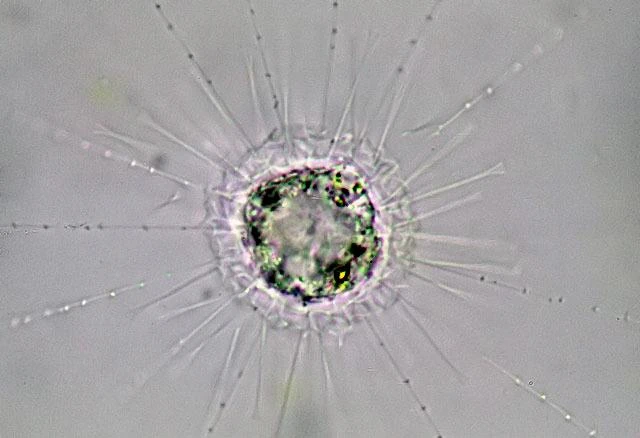
(上图:$R.~tubifera$)
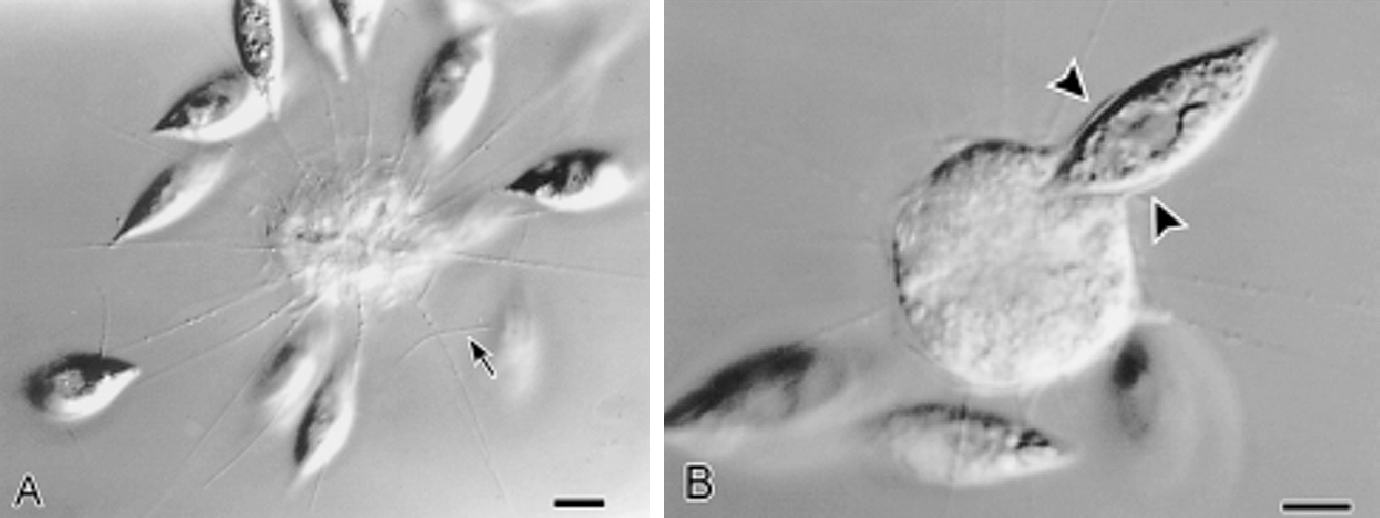
此外,刺囊虫的轴足在机械扰动等刺激下能快速收缩,并在几分钟内恢复;在其他中阳虫中也能观察到类似的现象。此过程中,微管束系统经历了快速的拆卸和(较慢的)重建,而我们目前还难以理解它们具体是如何做到的。
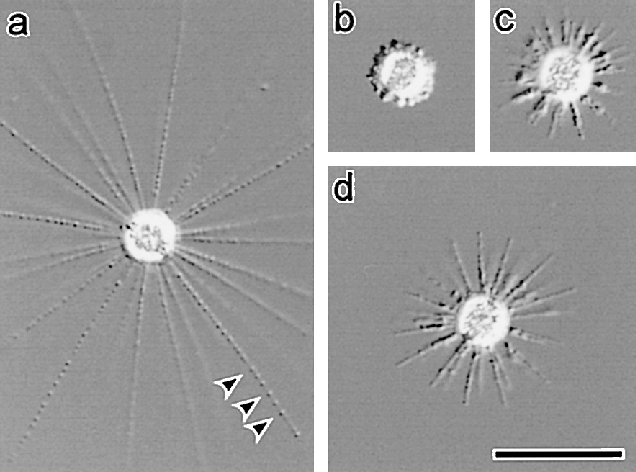
(上图:$R.~contractilis$ 的轴足收缩后恢复($b\to c\to d\to a$))
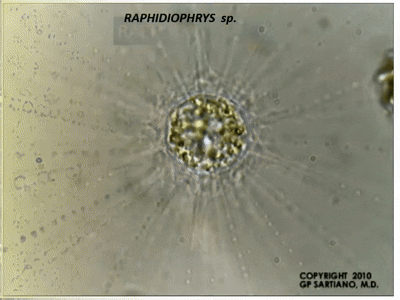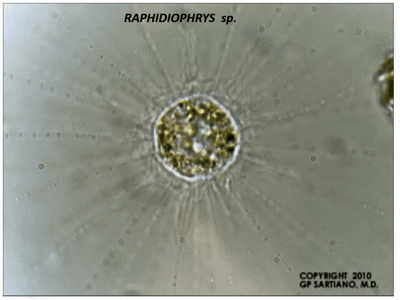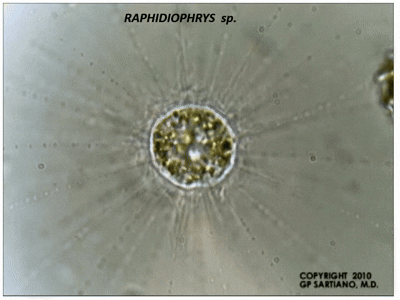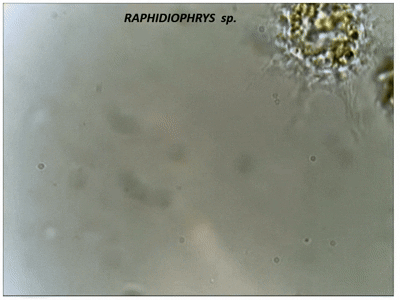
(上动图:$R.~contractilis$ 轴足快速收缩的珍贵影像。该物种在很长一段时间内都被错误地认为是刺日虫,因此图中有错误的标注)
当然,属内也有一些长得比较奇怪的。比如 $R.~pallida$,它们的鳞片层在某些地方强烈弯曲,形成许多根粗大的“刺”,像是毛笔的笔尖;整体来看有点像是六芒星,虽然它肯定不止“六芒”:
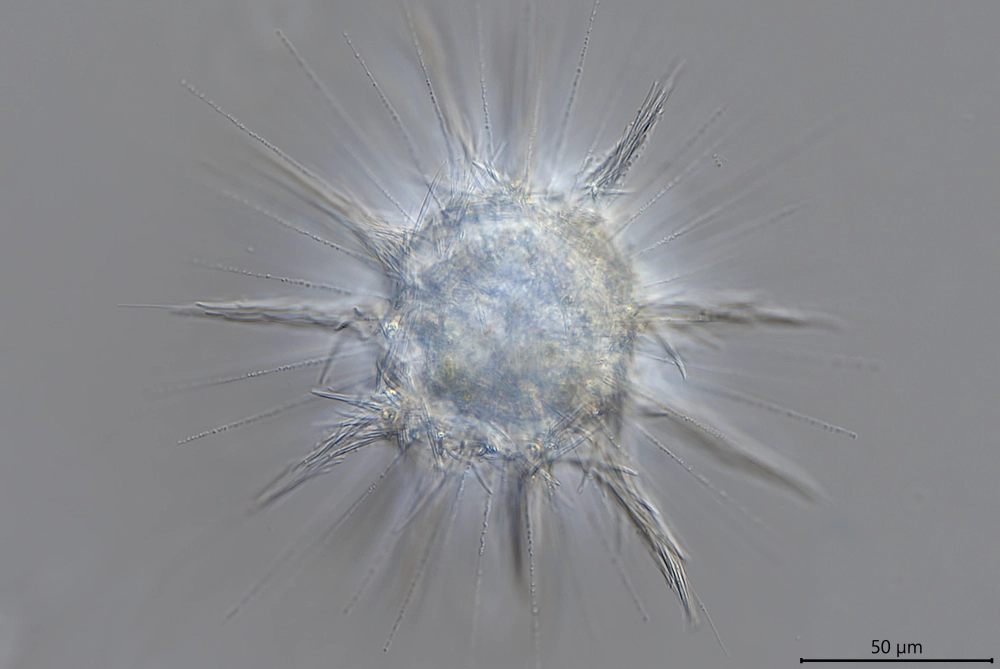
(上图:$R.~pallida$)
如果是刺囊虫是假货,那假刺囊虫($Pseudoraphidocystis$)就是假货中的假货了。该属成员都非常迷你,直径在 $10μm$ 左右,最大的特征是具有周质体,且有可能与棘胞虫的类似结构同源。因此,虽然没有分子生物学证据,但此处还是将其归入了棘胞虫科($\text{Acanthocystidae}$),作为棘胞虫属的姐妹。
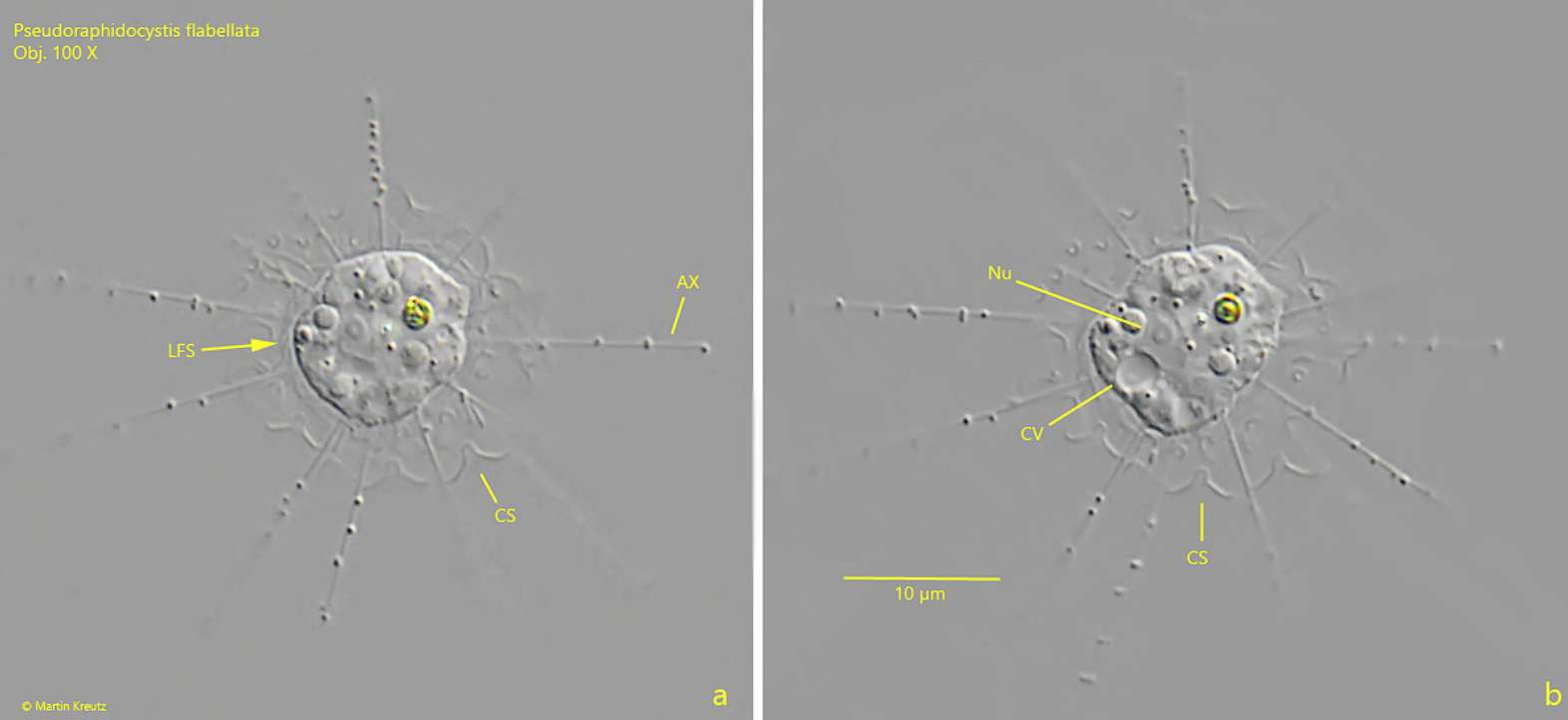
(上图:$P.~flabellata$)
海日虫($Marophrys$)长得比较像异日虫,只有尖刺而没有鳞片,不同之处在于海日虫没有明显的黏液套,且直径仅有 $10μm$。它们生活在海洋与咸水湖中,胞内经常可以见到共生的绿藻;进一步的研究发现,海日虫的线粒体内已经出现了与绿藻十分相似的基因序列,表明了基因转移事件的发生。合理的推测是,这些绿藻正在从临时的内共生体向真正的细胞器演变。
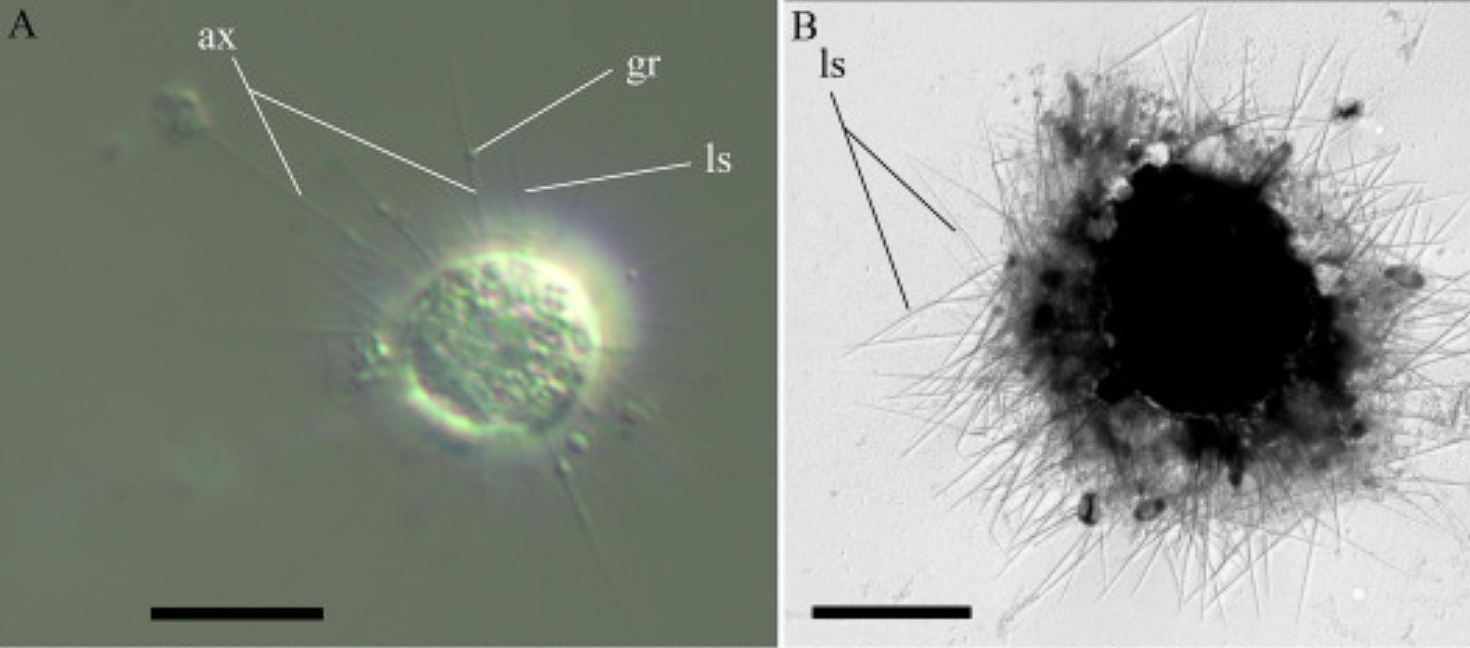
(上图:$M.~nikolaevi$ 的活细胞(左)与风干细胞(右))
瑞克虫($Ricksol$)则是棘胞虫目($\text{Acanthocystida}$)的基干类群。它们的尖锐鳞片在细胞外排布得无比杂乱,已经到了令人见之胆寒的地步。在超微结构分析中,还发现这些鳞片表面布满了各种各样的突起,可能起到提升摩擦力,以使鳞片层不会散架的作用:

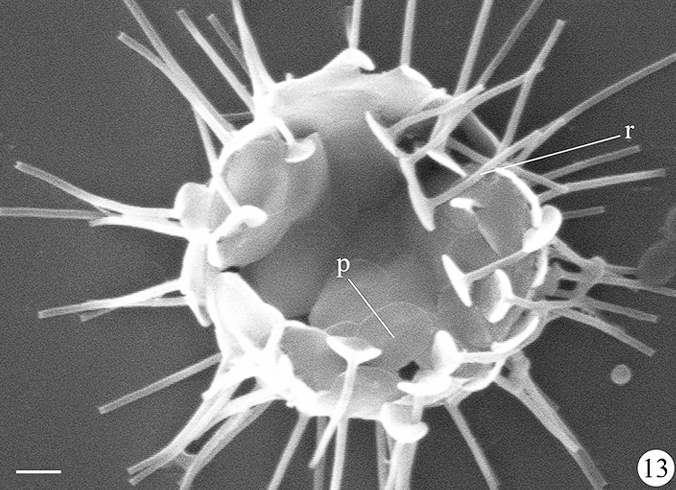
(上图:$R.~blepharistes$)
古棘胞虫目($\text{Chthonida}$)是泛棘胞虫纲的另一基干分支,棘胞虫目的姐妹。目下仅有两属,其中较晚发现的皮纳塔虫($Pinjata$)有两种扁平的鳞片,一种紧密地贴着细胞,另一种则松散地堆在外面,并在轴足伸出的地方上翘。其名字来源于一种传统玩具 $\text{Piñata}$,虽然我是看不出两者有什么相似:
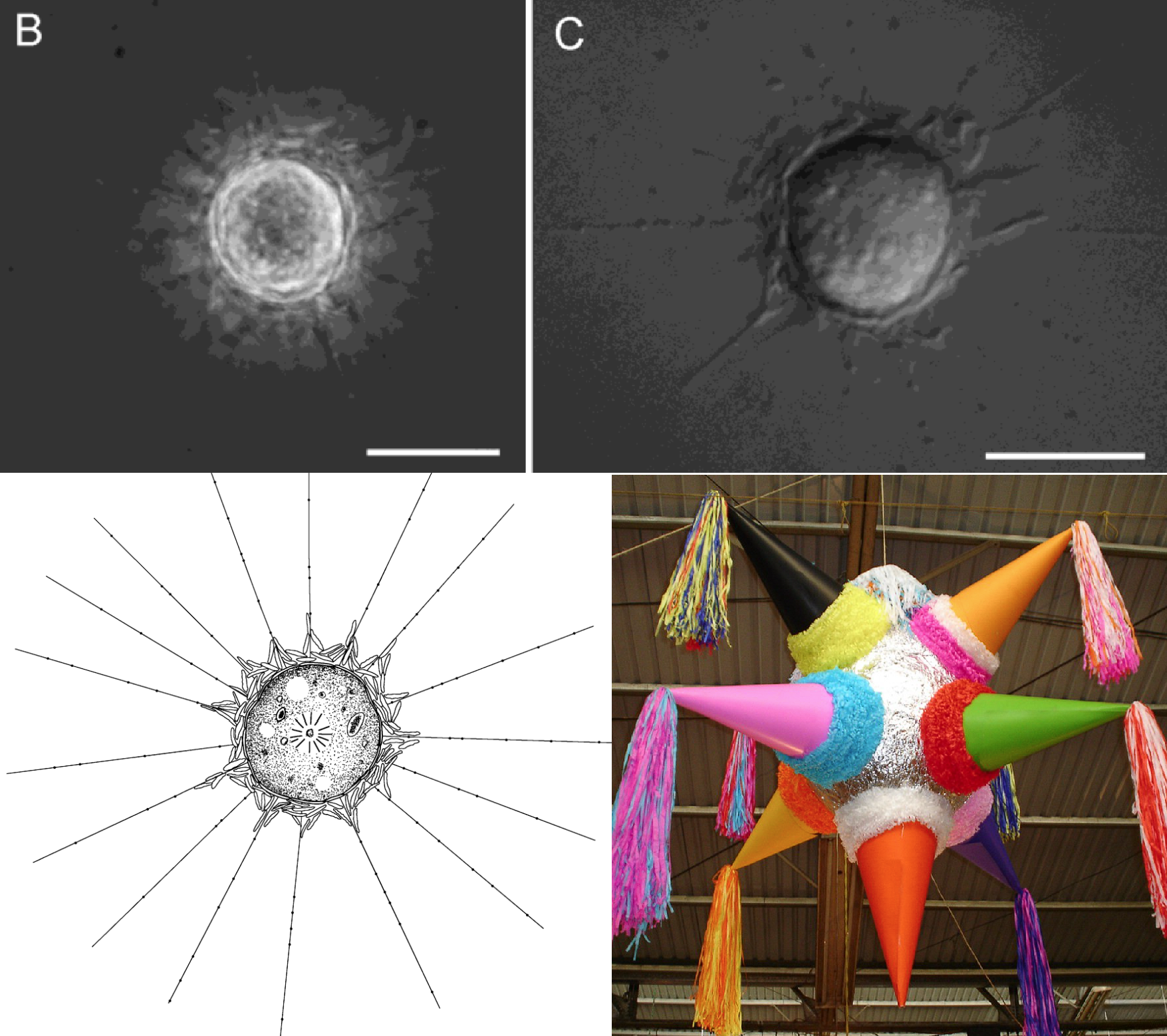
(上图上:$P.~ruminata$ 活细胞;左下:$P.~ruminata$ 简笔画;右下:那种叫 $\text{Piñata}$ 的玩具(这真的像吗?我觉得倒是上面那个 $R.~pallida$ 与它更像))
而另一个犹格虫属($Yogsothoth$),就值得我们大书特书了。在上文中,我们看到很多中阳虫族裔都独立演化出了群落,犹格虫也是如此。但是,犹格虫在这方面显然走得更远,以至于用“群落”来形容已经有些不太恰当。激进的观点认为,犹格虫已经演化为了真正的多细胞生物。
首先,它们的多细胞集体已经有了显著的细胞分化:外围细胞的轴足极长,可以达到细胞直径的十几倍;而内部细胞的轴足已经完全退化,不留痕迹。虽然没有进一步的证据,但犹格虫的内外细胞很有可能已经完成了分工,一个负责营养,另一个则负责繁殖等事务。其次,它们的多细胞集体已经有了固定的形态,所有细胞都紧密地贴在一起,还有一层非常厚的外鳞片将整个集体包裹。同时,每个细胞表面还有一层内鳞片,这显然是祖征,也表明它们不可能是合胞体。
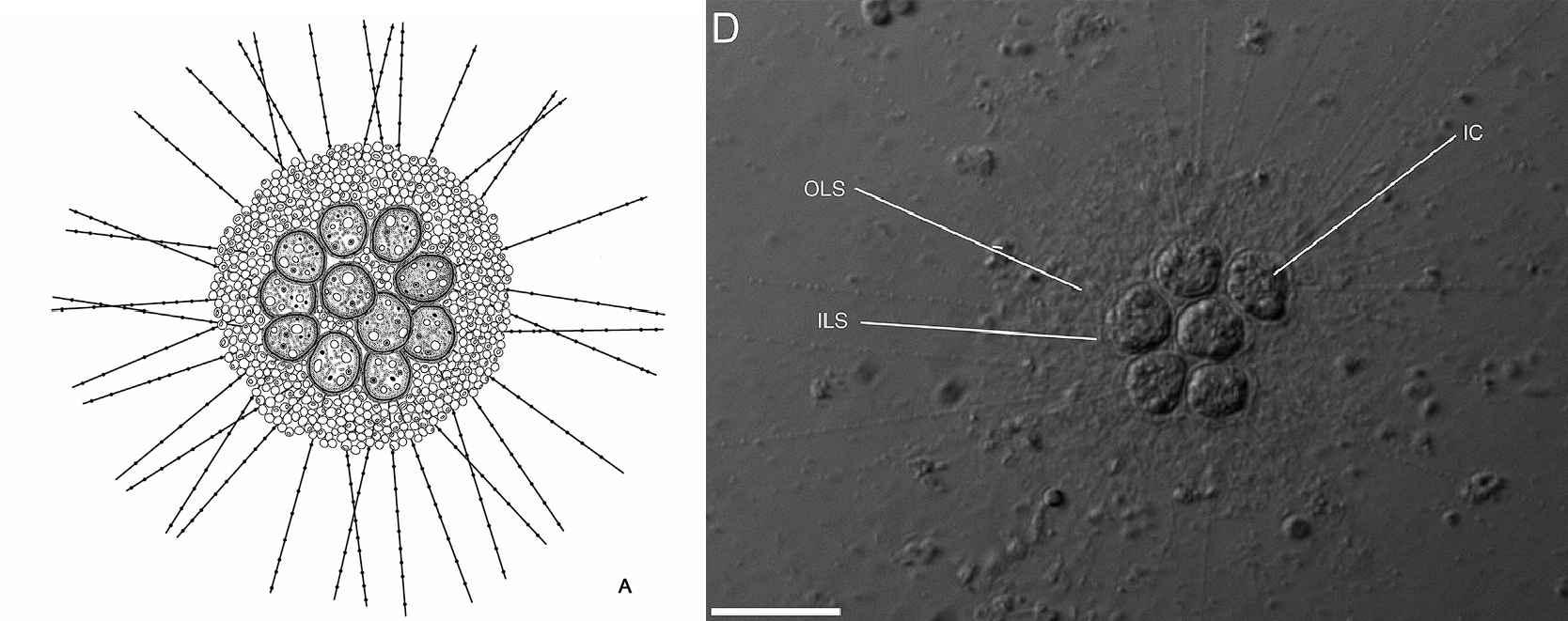
(上图:$Y.~knorrus$ 多细胞集体的简笔画与真实影像)
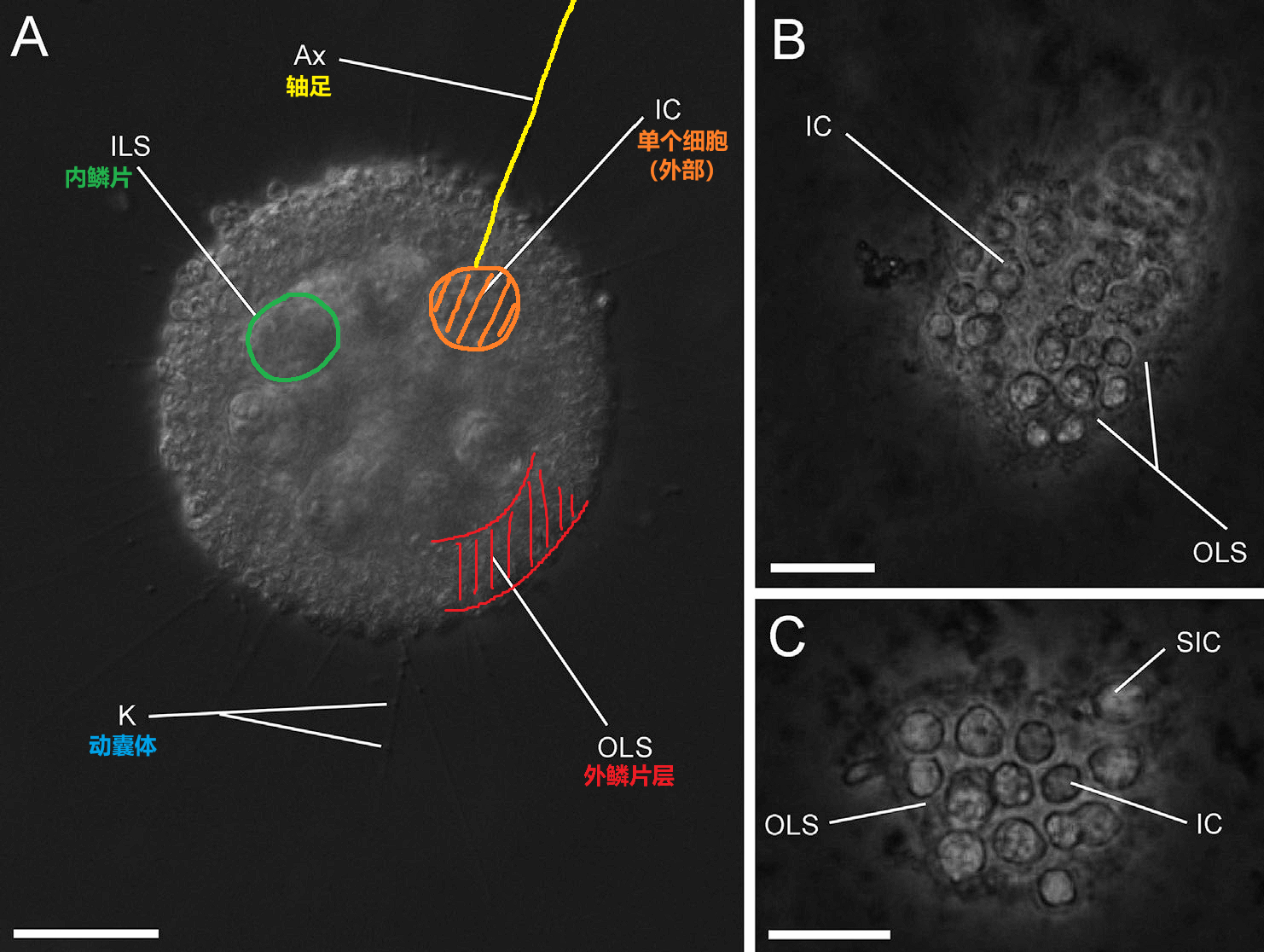
(上图:较大的犹格虫集体。图 $\text{A}$ 中标出了上文所述的各种结构)
此外,犹格虫集体可以直接进行分裂生殖,即较大的母集体从中间裂开,形成两个子集体;二者各占有原来外鳞片层的一般,并在此后的生长中独立造出另一半。它们有时也会产生单个的细胞(可能还会给它附带一些营养物质),但这个细胞很快就会在一系列分裂后变成一个新的集体。
根据外鳞片微结构,犹格虫被识别为两个物种。$Y.~knorrus$ 的单个鳞片像个椰子壳,而 $Y.~carteri$ 的则像个壶,开口更小且空腔更深。鳞片表面有微小的突起,与前面讲到的瑞克虫类似,可能也是为了提升鳞片之间的摩擦力:

(上图上半部分:$Y.~knorrus$ 的外鳞片;下半部分:$Y.~carteri$ 的外鳞片)
最令人惊奇的是,它们居然可以主动运动,虽然只是简单且缓慢的蠕动,但在中阳虫中也是独一无二了。我们至今仍难以理解它们是如何在披着如此厚重的鳞片的情况下用那纤细的轴足进行运动的,但这显然说明犹格虫细胞之间的配合已经达到了很高的境界。
那么,它们真的已经完成了由单细胞到多细胞的蜕变了吗?很难说。“多细胞”与“单细胞群落”两个词之间本就没有明显的界限,所以遇到犹格虫这样卡在二者之间的,自然是公说公有理,婆说婆有理。那就把这个问题留给各位读者吧,相信看到这里的你,心中也已有了自己的答案。
3. 关于分类学的讨论
在上文中,我们已经讨论了中阳虫门中所有已发现的属(截至我写这句话的时候)。但是,我们并没有过多地讨论分类学,因为小小一个中阳虫门,内部分类直到今天仍然混乱,所有我刻意把这部分放在了后面。这一部分十分枯燥,所以若你对分类学没有兴趣,大可直接跳过此章节。
首先先说外部演化位置。传统上,定鞭界($\text{Haptista}$)只有两门,即中阳虫门与定鞭藻门($\text{Haptophyta}$),二者显然互为姐妹群,且都是基干分支;但是 $2024$ 年的一项分子生物学研究把另一个名为网鞭虫门($\text{Telonemia}$)的类群也拉了进来,且认为它是定鞭藻的姐妹,如下图:
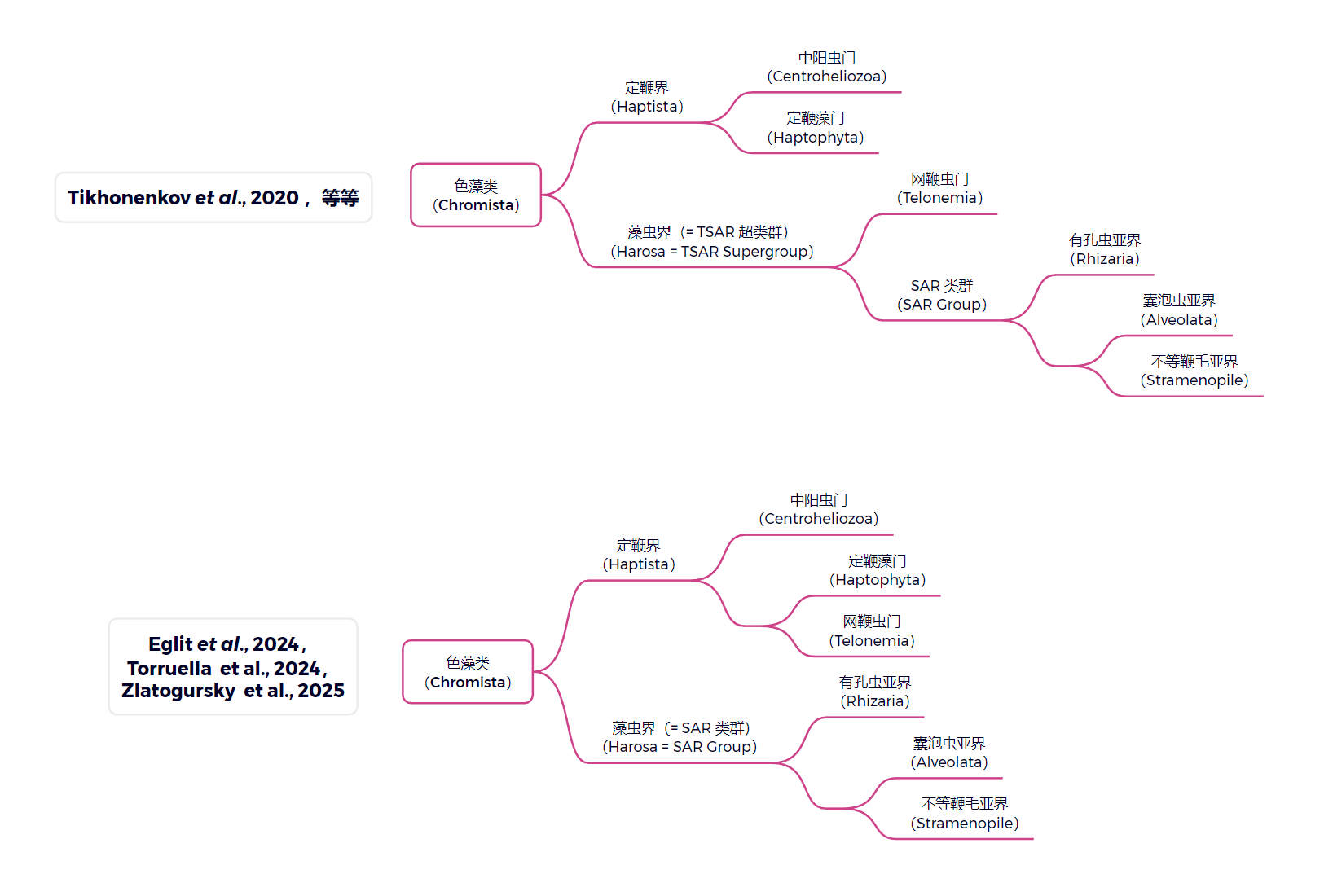
(一篇关于网鞭虫的文章:Link)
这样一来,中阳虫不再是定鞭藻的姐妹,且知名的 $\text{TSAR}$ 超类群也就没了,藻虫界($\text{Harosa}$)与 $\text{SAR}$ 类群成为了同义词。目前看到的已经有 $\text{3-4}$ 篇研究支持这一观点了,所以我也在逐渐接纳这种说法。
中阳虫门内部争议也很大。首先是基部难以确定,但因为很多大类群都这样,所以也见怪不怪了。其次是关于翼阳虫的问题,前面也提到,翼阳虫属不是单系群,理应被拆分,但这已经是各路研究唯一的共识点了。至于它具体是怎么样的,当然是莫衷一是,有人说要拆成 $5$ 类,也有人说 $3$ 类就够,当然也有人说更多。另一个非单系的属唤作 $Polyplacocystis$ ,它在 $2018$ 年即被废止,其下物种均根据分子证据并入刺日虫属与刺囊虫属。
随后,还有关于幽灵类群的问题。如果你仔细地看了每张系统发育图的话,会注意到一个叫异刺日虫属($Heteroraphidiophrys$)的类群被打上了问号。事实上,该类群从未被正式发布,其下包含唯一物种 $H.~australis$,最早是在一篇非公开的文献中有提及,后来被 $Mikrjukov$ 写进了他的专著中从而为人所知。$2015$ 年,$Plotnikov$ 又称在俄罗斯西南部的河流中发现了异刺日虫,但未拍摄活细胞图片,也没有进一步培养。总之根据 $\text{ICZN}$ 的规则,该名称其实是无效的。$2019$ 年,皮纳塔虫($Pinjata$)的发现者注意到二者的相似之处,怀疑所谓 $H.~australis$ 其实属于皮纳塔虫属。
在正式发布的类群中,上文已经提到了神秘的副球星虫($Parasphaerastrum$)。事实上,该属根本没有分子证据,系统发育位置只是根据形态。另外,$2022$ 年的一篇研究怀疑副球星虫其实与刺囊虫($Raphidocystis$)有关,并直接指出它可能是 $R.~pallida$(不记得这货长什么样的可以去翻一翻)的一支更加特化的近亲。这种分析基于他们对刺囊虫属的系统发育重建,是有一定道理的:
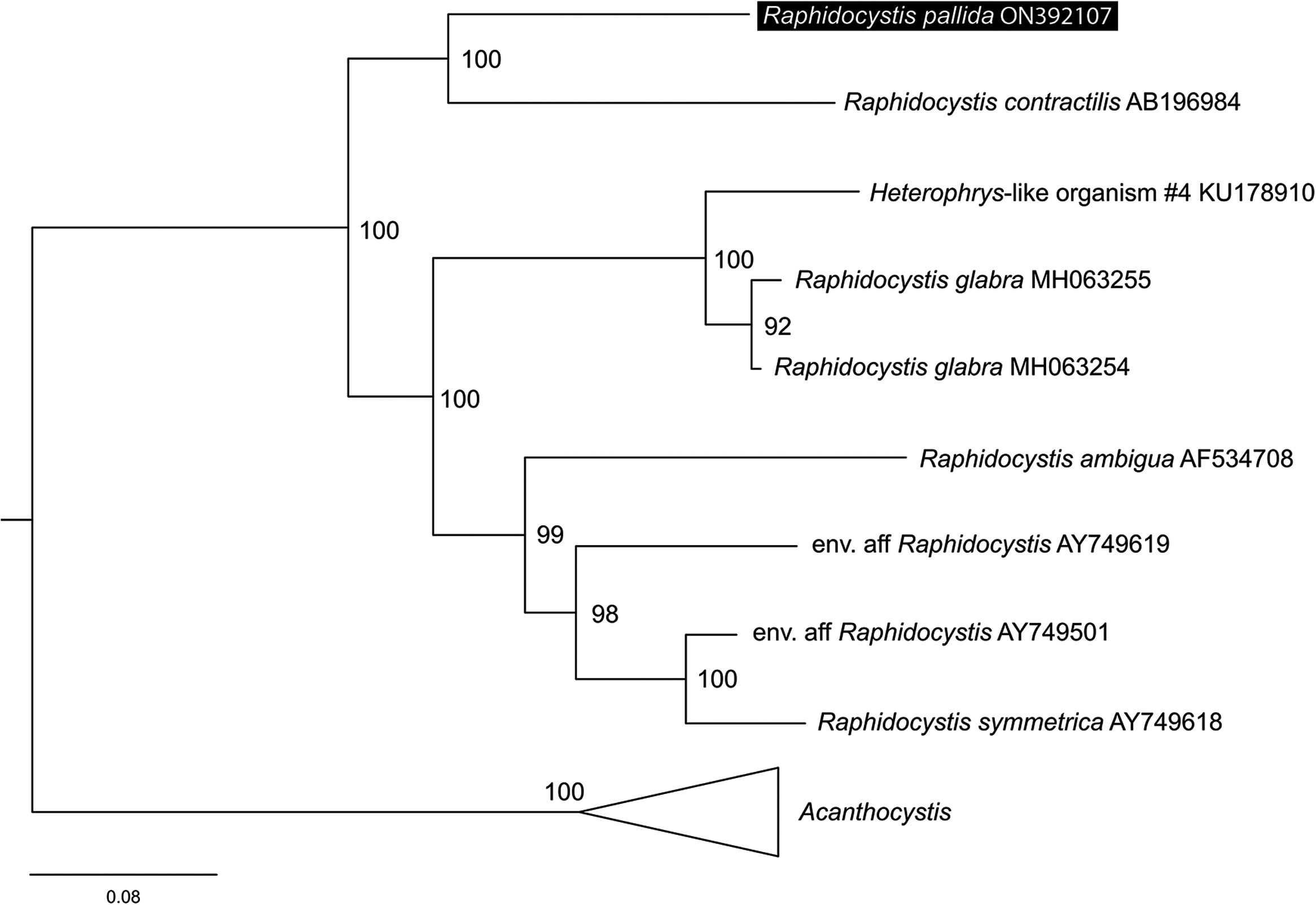
(上图:刺囊虫属($Raphidocystis$)内部各物种的系统发育)
自从发现了一些中阳虫具有二态性之后,更多物种被推上了怀疑的舞台。由于已发现的具有二态形的类群彼此之间关系极远,所以合理推测二态形其实在中阳虫中是普遍的。$2020$ 年,就有一种未正式发表的“类异日虫生物”被证明是一种异日虫 $R.~heterophryoidea$ 的一个生命阶段。
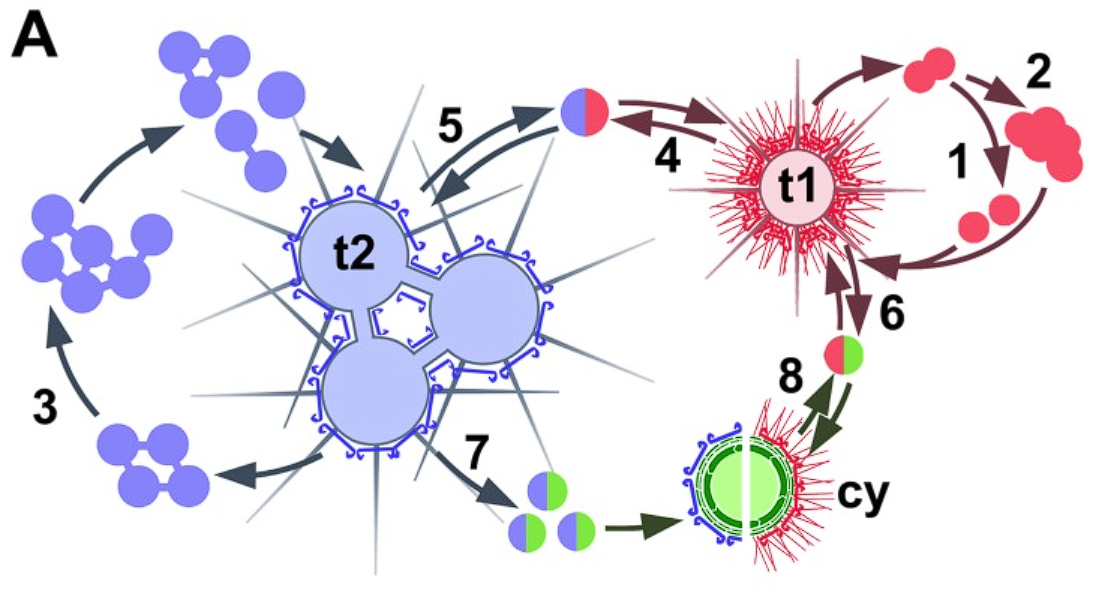
(上图:$R.~heterophryoidea$ 的生命周期示意,可见在 $\text{t1}$ 阶段表现出异日虫的特征,无鳞片而有密集的尖刺)
另外,中阳虫中存在广泛的趋同演化,最经典的便是刺日虫和刺囊虫。$2018$ 年,$Zlatogursky$ 等人将 $7$ 个刺日虫物种重新确定为刺囊虫,名单如下:

直到我写这句话的这一刻,维基百科等网站依然没有更改,所以如果你想要深入了解的话,还请自行阅读相关文献。
另外,关于领囊虫属($Choanocystis$)也有一些需要补充,那就是该类群现在的正式名其实应该是 $Ozanamia$,但事实上,该名称未被广泛使用,毕竟 $Choanocystis$ 这个名字已经用了一百多年了。至于为什么要改,是因为 $Penard,1904$ 用模式种 $C.~lepidula$ 建立该属时没有将其放入中阳虫门。有两种可能,其一是 $Penard$ 归类时的错误,其二是我们现在认为的 $C.~lepidula$ 和 $Penard$ 最初发表的根本不是一个东西。总之,$Choanocystis$ 这个名字确实在规则上不太过得去,但我也不赞成直接改名,毕竟大家都用惯了,你一纸论文就想改过来也不太行,所以上文中也没有改。
其实,关于中阳虫的分类还有很多可以碎碎念的,但我实在不想写下去了,那就这样吧。
4. 结语
如你所见,这是一类极不知名的生物,但就我个人而言,还是更加喜好这种冷门的生物类群。为了了解它们,我必须耗费更多的精力去查阅文献、苦思冥想,或许直到精疲力尽才能整理出这么一篇不起眼的文章。但是,我在提笔之前便已做好准备,而当我写到这里时,就仿佛是在无数天筚路蓝缕之后终于到达了这片未知之境的边缘,那种快乐难以言表;而我总是希望,各位读者可以与我分享这份喜悦。
在最后,贴上一张完整的,中阳虫从门到属的系统发育树:
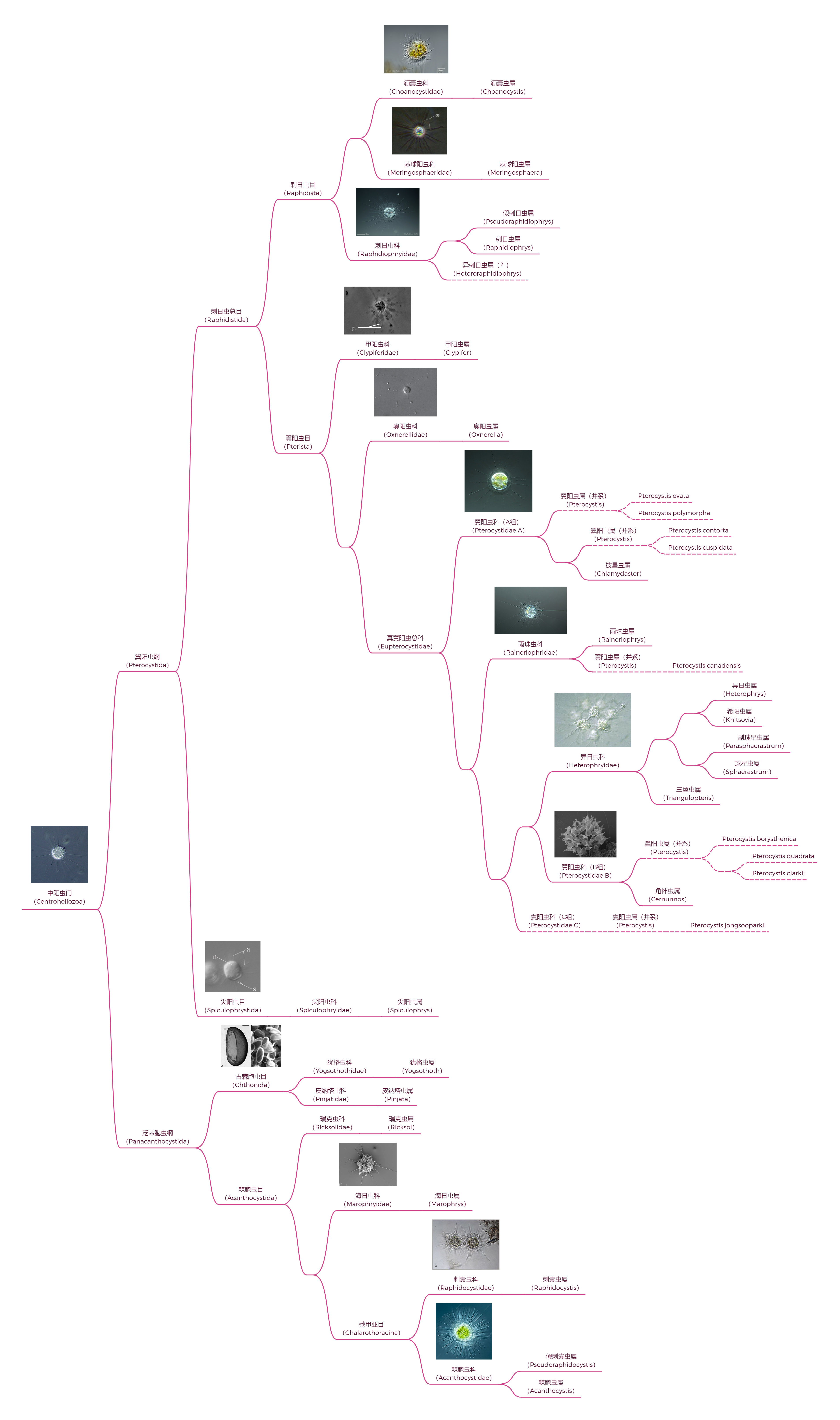
(好吧,图片依然不清楚)以及它的 $\text{pdf}$ 版链接:Link
5.参考资料
网站
$[1]$:Siemensma, F. J., Microworld, world of amoeboid organisms. World-wide electronic publication, Kortenhoef, the Netherlands. Searched on May 3, 2025. Link(这是一个非常不错的网站,收集了上万张伪足虫的图片)
书籍
$[2]$:Handbook of the Protists (Second Edition), John M. Archibald, Alastair G.B. Simpson, Claudio H. Slamovits, 2017 Link
论文
$[3]$:Zlatogursky, V.V., Shɨshkin, Y., Drachko, D. & Burki, F. (2021) The long- time orphan protist Meringosphaera mediterranea Lohmann, 1902 [1903] is a centrohelid heliozoan. Journal of Eukaryotic Microbiology, 68, e12860 Link
$[4]$:Keeling PJ, Eglit Y (2023) Openly available illustrations as tools to describe eukaryotic microbial diversity. PLoS Biol 21(11):e3002395. Link
$[5]$:Sakaguchi M, Suzaki T, Khan S M M K, et al. Food capture by kinetocysts in the heliozoon Raphidiophrys contractilis[J]. European journal of protistology, 2002, 37(4): 453-458. Link
$[6]$:Yabuki A, Chao E E, Ishida K I, et al. Microheliella maris (Microhelida ord. n.), an ultrastructurally highly distinctive new axopodial protist species and genus, and the unity of phylum Heliozoa[J]. Protist, 2012, 163(3): 356-388. Link
$[7]$:Bardele C F. Comparative study of axopodial microtubule patterns and possible mechanisms of pattern control in the centrohelidian heliozoa Acanthocystis, Raphidiophrys and Heterophrys[J]. Journal of Cell Science, 1977, 25(1): 205-232.Link
$[8]$:Zlatogursky V V. There and back again: parallel evolution of cell coverings in centrohelid heliozoans[J]. Protist, 2016, 167(1): 51-66. Link
$[9]$:Zagumyonnyi D G, Tikhonenkov D V. A new centrohelid heliozoan, Pterocystis polycristalepis sp. nov., and taxonomic and phylogenetic concerns within Pterista (Haptista: Centroplasthelida)[J]. European Journal of Protistology, 2024, 94: 126064. Link
$[10]$:Zlatogursky V V, Drachko D, Klimov V I, et al. On the phylogenetic position of the genus Raphidocystis (Haptista: Centroplasthelida) with notes on the dimorphism in centrohelid life cycle[J]. European Journal of Protistology, 2018, 64: 82-90. Link
$[11]$:Zlatogursky V V, Gerasimova E A, Drachko D, et al. Pinjata ruminata gen. et sp. n.—A new member of centrohelid family Yogsothothidae (Haptista: Centroplasthelida) from the brackish river[J]. Journal of Eukaryotic Microbiology, 2019, 66(6): 862-868. Link
$[12]$:Khan S M M K, Arikawa M, Omura G, et al. Axopodial contraction in the heliozoon Raphidiophrys contractilis requires extracellular Ca2[J]. Zoological science, 2003, 20(11): 1367-1372. Link
$[13]$:Zlatogursky V V. Three new freshwater species of centrohelid heliozoans: Acanthocystis crescenta sp. nov., A. kirilli sp. nov., and Choanocystis minima sp. nov[J]. European journal of protistology, 2010, 46(3): 159-163. Link
$[14]$:Shishkin Y, Drachko D, Zlatogursky V V. Clypifer cribrifer gen. nov., sp. nov.(Clypiferidae fam. nov., Pterocystida, Centroplasthelida), with notes on evolution of centrohelid siliceous coverings[J]. International Journal of Systematic and Evolutionary Microbiology, 2021, 71(7): 004856. Link
$[15]$:Cavalier-Smith T, Chao E E. Oxnerella micra sp. n.(Oxnerellidae fam. n.), a tiny naked centrohelid, and the diversity and evolution of heliozoa[J]. Protist, 2012, 163(4): 574-601. Link
$[16]$:Zagumyonnyi D G, Radaykina L V, Tikhonenkov D V. Triangulopteris lacunata gen. et sp. nov.(Centroplasthelida), a New Centrohelid Heliozoan from Soil[J]. Diversity, 2021, 13(12): 658. Link
$[17]$:Mikrjukov K A. Revision of Genera and Species Composition of Lower Centroheliozoa. II. Family Raphidiophryidae n. tam[J]. Archiv für Protistenkunde, 1996, 147(2): 205-212. Link
$[18]$:Zagumyonnyi D G, Radaykina L V, Keeling P J, et al. Centrohelid heliozoans of Ukraine with a description of a new genus and species (Haptista: Centroplasthelida)[J]. European Journal of Protistology, 2022, 86: 125916. Link
$[19]$:Shɨshkin Y, Drachko D, Klimov V I, et al. Yogsothoth knorrus gen. n., sp. n. and Y. carteri sp. n.(Yogsothothidae fam. n., Haptista, Centroplasthelida), with notes on evolution and systematics of centrohelids[J]. Protist, 2018, 169(5): 682-696. Link
$[20]$:Shɨshkin-Skarð Y, Drachko D, Zlatogursky V V. Shedding Light on the Origin of Acanthocystidae: Ricksol blepharistes gen. n., sp. n.(Ricksolidae fam. n., Panacanthocystida, Centroplasthelida), with Notes on the Evolution of the Genera Acanthocystis, Ozanamia gen. n.(Ozanamiidae fam. n.), and “Heterophrys-like Organisms”[J]. Organisms Diversity & Evolution, 2023, 23(2): 263-274. Link
$[21]$:Cavalier-Smith T, von der Heyden S. Molecular phylogeny, scale evolution and taxonomy of centrohelid heliozoa[J]. Molecular Phylogenetics and Evolution, 2007, 44(3): 1186-1203. Link
$[22]$:Nishimura Y, Shiratori T, Ishida K, et al. Horizontally-acquired genetic elements in the mitochondrial genome of a centrohelid Marophrys sp. SRT127[J]. Scientific Reports, 2019, 9(1): 4850. Link
$[23]$:Gerasimova E A, Radaykina L V, Zagumyonnyi D G, et al. Morphology and spicules elemental composition of Marophrys nikolaevi spec. nov.(Haptista: Centroplasthelida)[J]. European Journal of Protistology, 2022, 84: 125888. Link
$[24]$:Drachko D, Shɨshkin Y, Zlatogursky V V. On the phylogenetic position of Raphidocystis pallida with some notes on its life cycle[J]. Journal of Eukaryotic Microbiology, 2022, 69(4): e12916. Link
$[25]$:Gerasimova E A, Mindolina Y V, Tikhonenkov D V, et al. Unexpected ubiquity of heart‐shaped scale morphotype in Centroplasthelida (Haptista): Ancestral trait or multiple acquisitions?[J]. Journal of Eukaryotic Microbiology, 2023, 70(6): e12992. Link