1. 引言
如果你研究过真核生物的早期演化,就一定会注意到这么一类生物:后滴虫($\text{Metamonads}$)。它真可谓是真核生物中最神秘的类群之一,科学家们为它的身世打了十几年的嘴炮,却依然没有达成共识。但大家都承认,后滴虫的系统发育位置研究极为重要。
之所以这么说,是因为我们相信在这个远古幽灵的身上,潜藏着解开真核生物早期演化之谜的钥匙。
(上图:披发虫($Trichonympha$),属副基体门披发虫纲,是许多蜚蠊目昆虫的肠道共生虫,帮助它们消化纤维素)
(上图:副三鞭虫($Paratrimastix$),属前轴柱门副三鞭虫目,自由生活于淡水淤泥之中)
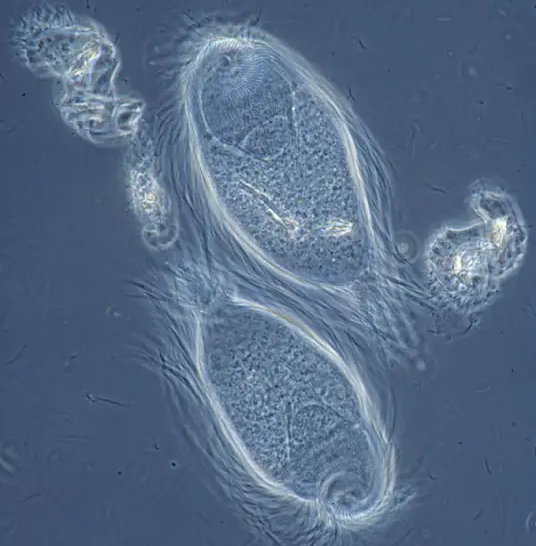
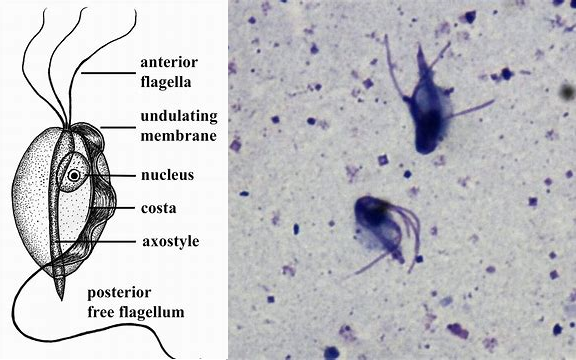
(上图:阴道滴虫($T.~vaginalis$),著名的人类寄生虫,依靠性行为传播,属副基体门毛滴虫纲毛滴虫属)
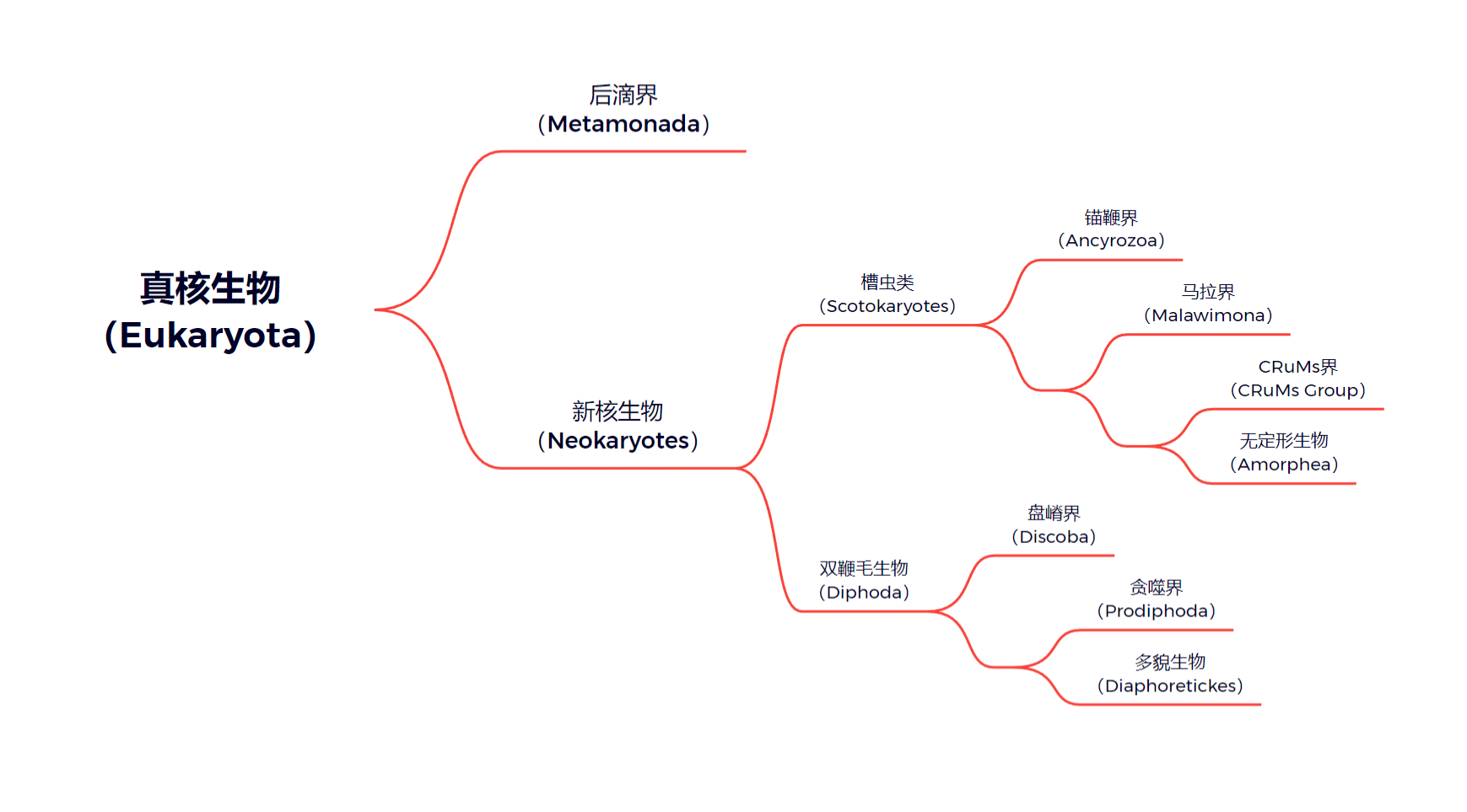
早在十七世纪,一些寄生性的后滴虫就已被人类发现。在很长的时间里,人们只知道它们是真核生物,后来又被扔进了原生生物这个大筐中。2002 年,著名系统发育学家 $\text{T. Cavalier-Smith}$ 建立了古虫界($\text{Excavata}$,请勿与脊索动物门中的“古虫”混淆),后滴虫成为了其中的一门。只是随着分子生物学的发展,人们意识到所谓古虫并不是单系群,因此将其从正式分类群中除名,而后滴门也随之成为了后滴界($\text{Metamonada}$)。
后滴虫最大的特点,便是没有传统意义上的线粒体,取而代之的是氢化酶体($\text{hydrogenosome}$)、线裂体($\text{mitosome}$)等奇怪的细胞器,这也使得后滴虫全员厌氧。这些特征和所有分子生物学证据都指向一个结论——后滴虫位于真核生物演化树基部。争议的焦点在于其具体的演化位置,以及其是否是单系群。我们会在文章的末尾着重讨论这些争论,而下图是我个人支持的观点,即后滴虫是真核生物最基干的单系群:
那么现在,就让我们正式走进这个神秘的世界。
2. 后滴虫的内部演化
传统上,后滴虫有三大门:拱形虫门、副基体门和前轴柱门。$2017$ 年,厌氧变形虫被发现,并很快被确认为副基体门的姐妹;同样在近年被发现的巴塞尔虫($Barthelona$)和驼背虫($Skoliomonas$)构成了名为“$\text{BaSk}$”的单系群,是为拱形虫门的姐妹。目前认为的这五大类群间的关系如下图(K. Williams et al. 2024):
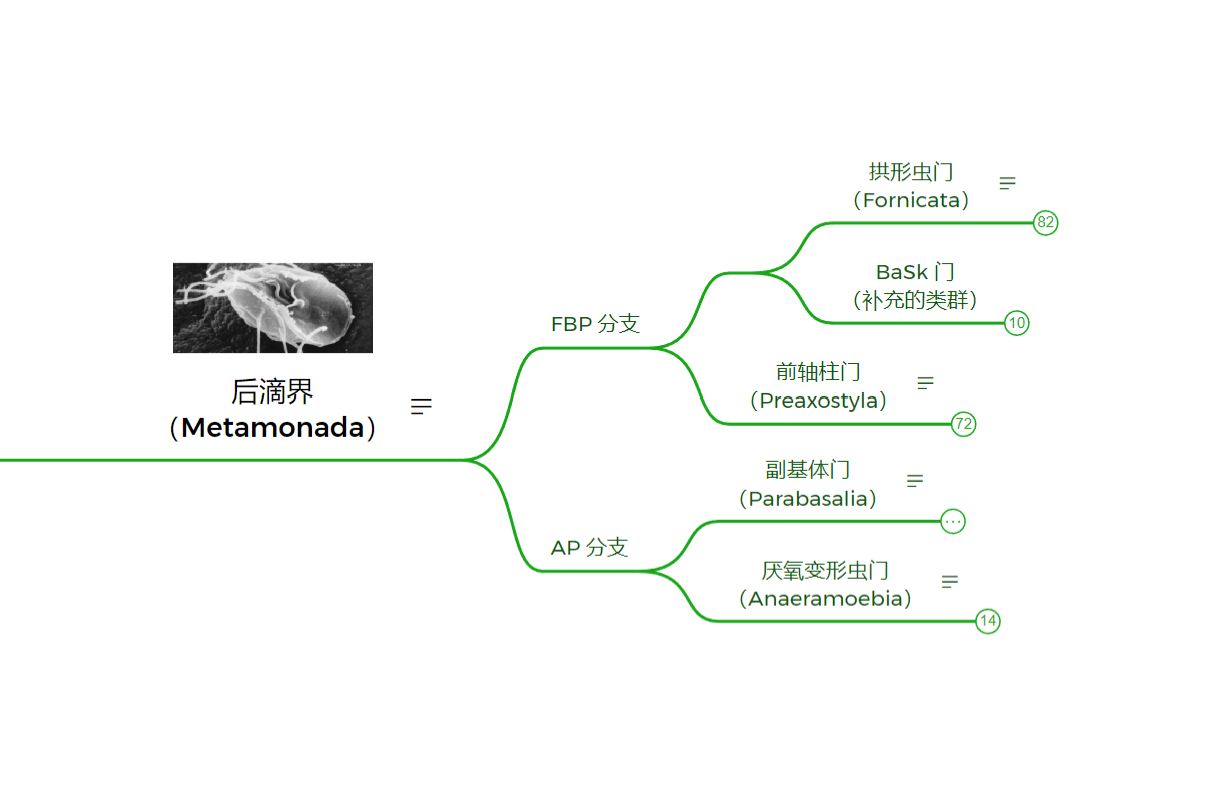
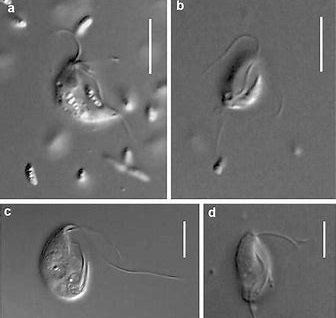
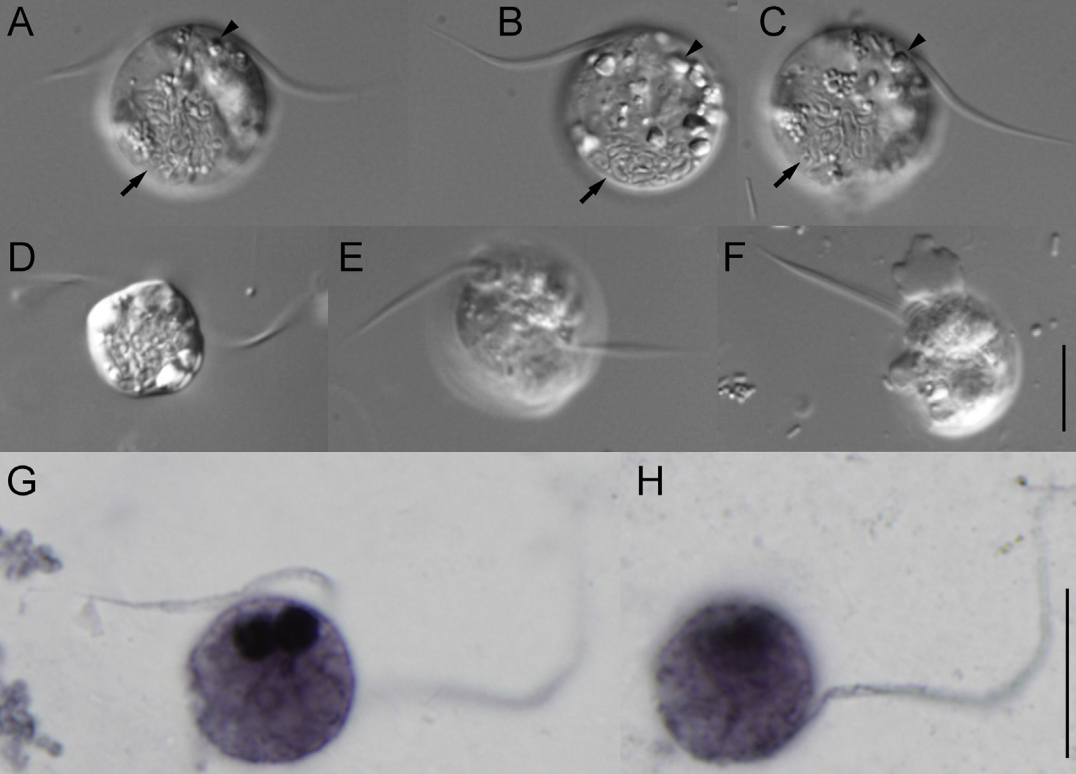
五大门中分支最少的是厌氧变形虫门($\text{Anaeramoebia}$),目前只发现了一个属。它们自由生活于世界各地缺氧的海洋沉积物中,可以伸出伪足用于运动,或捕食周遭的原核生物。除此之外,$A. ignava$ 还有还有一种鞭毛虫形态(较为罕见),呈现出圆形或梨形,可以快速运动。它似乎可以在两种形态间自由切换,以适应周围环境。
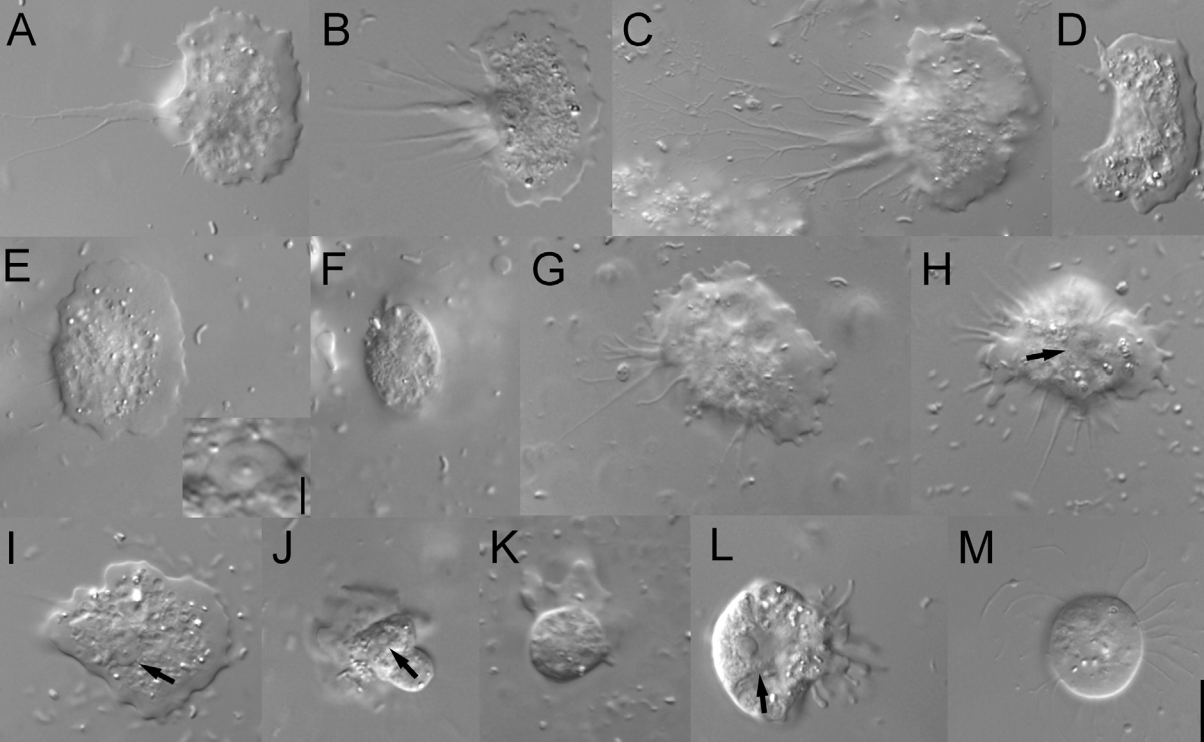
(上两图:变形虫和鞭毛虫形态的 $A.~ignava$)
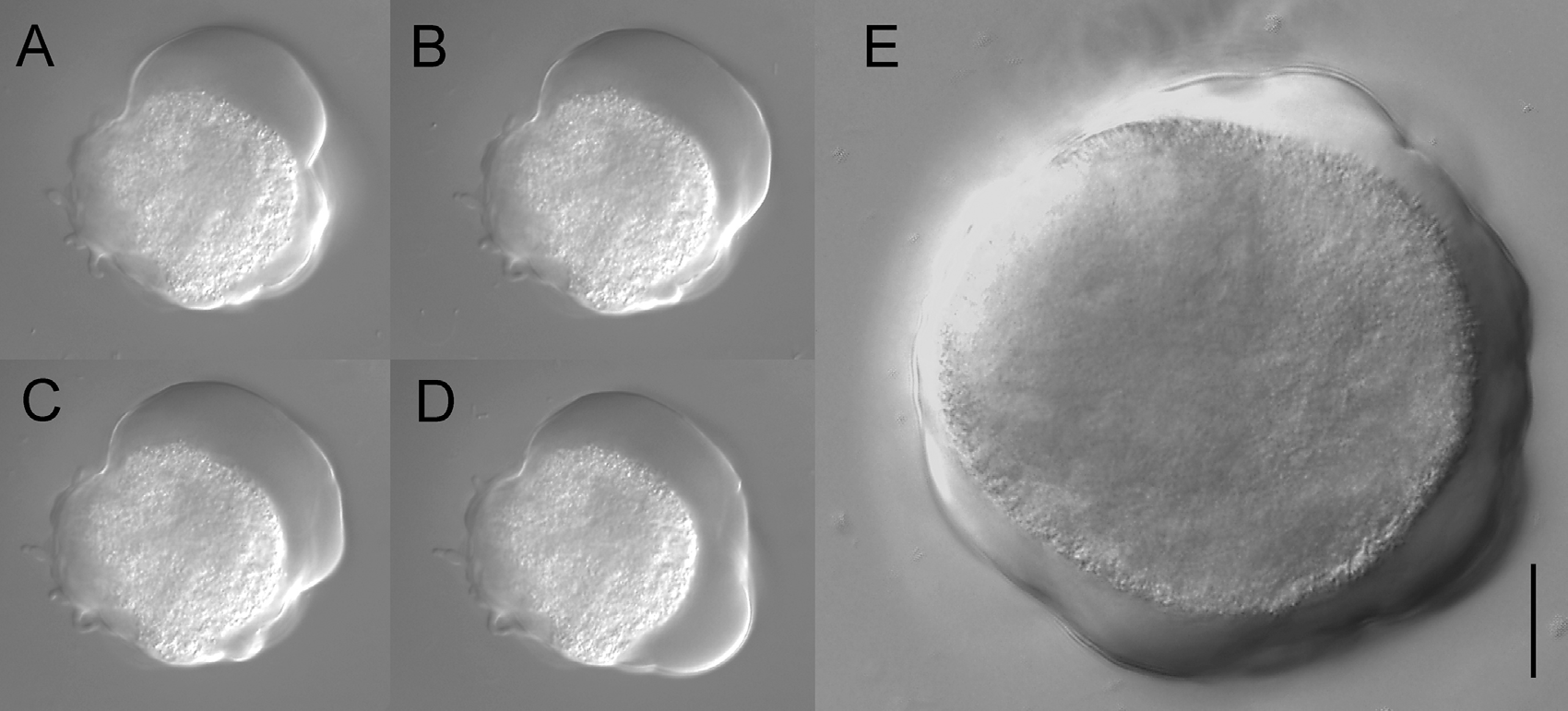
$A.~flamelloides$ 还可以形成与黏菌类似的原质团,比普通的变形虫和鞭毛虫大得多(可达 $0.1mm$ 以上),是具有多个细胞核的合胞体:
虽然厌氧变形虫分化很早,但在亿年又亿年的演化中,它们的面貌很可能已经与祖先大相径庭。目前认为,$A.~ignava$ 特有的鞭毛虫形态可能与祖先最为相近。恰好,多条分子生物学证据支持 $A.~ignava$ 的演化位置在整个属的最基部,与属内其他物种关系较远:

(上图:厌氧变形虫属内系统发育树,Táborsky et al. 2017)
与厌氧变形虫关系最近的副基体门($\text{Parabasalia}$)则是后滴界中最大的门级分类群。它们具有独特的细胞器:副基体($\text{parabasal body}$),位于高尔基体附近,可能起辅助后者工作的作用。传统上,副基体门内只有毛滴虫和披发虫两个类群,但 $2024$ 年的一项研究表明,该门内部的演化远比我们之前想象得复杂。重建的系统发育树将副基体门拆成了十一纲,如下图所示(Boscaro et al. 2024):
副基体门中大多数物种都营共生或寄生生活,且主要是在动物肠道和生殖道中。事实上,纵观所有生态位,肠道共生和寄生都是后滴虫的绝对优势,这与它们厌氧的习性有很大关联。但显然,它们也是由自由生活的祖先演化而来。
副基体门中最基干的宾巴维虫纲($\text{Pimpavickea}$)佐证了这一点。它们于 $2022$ 年被发现于水体沉积物中,最突出的特征便是在细胞前部拥有一个超长的可活动部分,被称为吻($\text{rostrum}$),具体有何作用尚不可知:
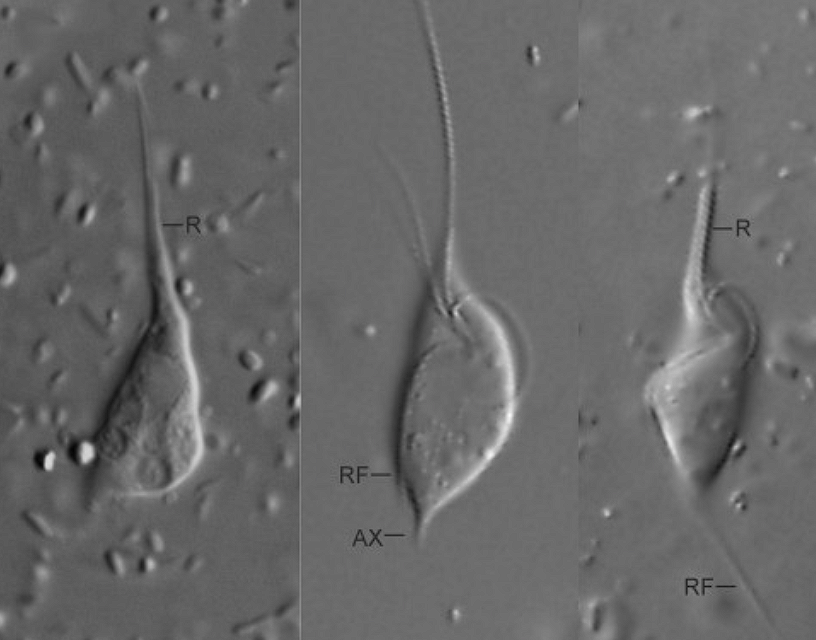
(上图:$P.~limacoides$)
除去宾巴维虫后,副基体门中剩下的物种可以分为两个大分支。其中,$\text{Cadamassta}$ 总纲被分子生物学强烈支持,虽然其下各类群在形态上少有共同点。该分支内尚未发现自由生活的物种,所有成员都共生或寄生于动物,尤其是昆虫和脊椎动物体内。
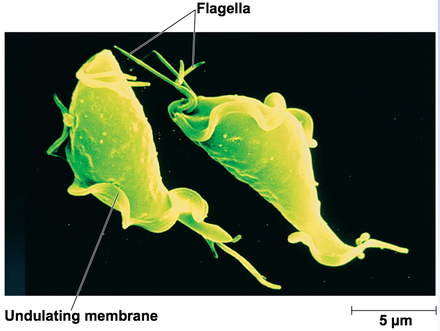
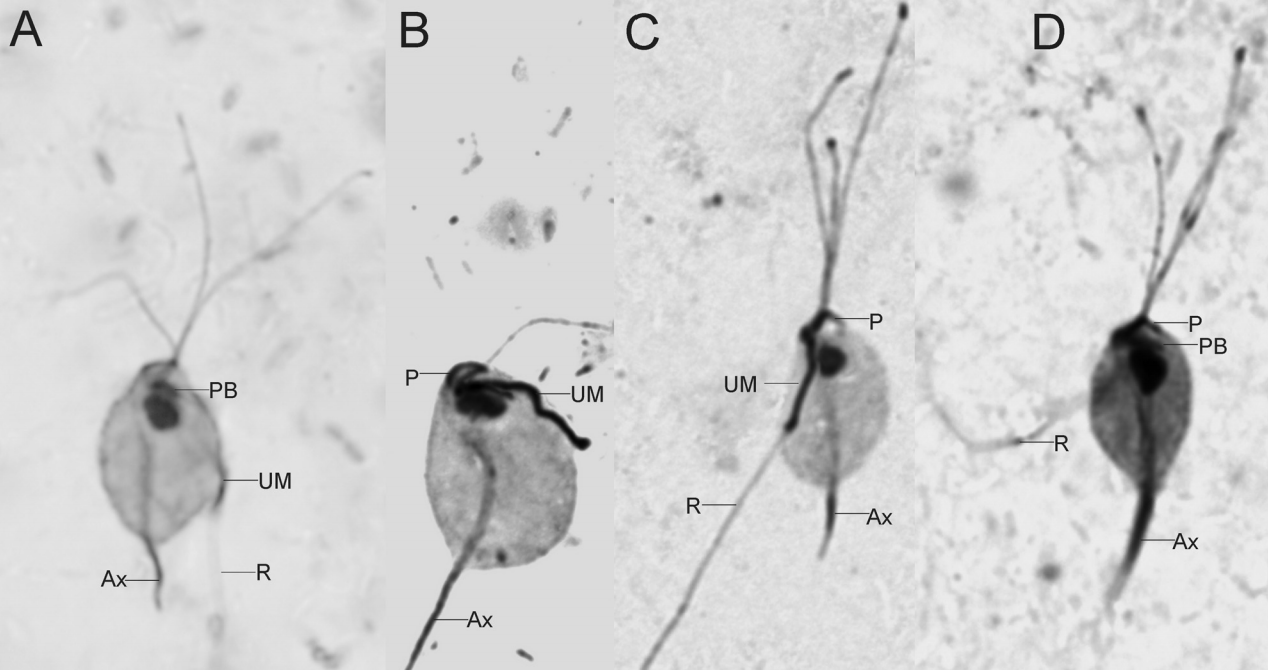
在这之中比较著名的如三毛滴虫纲($\text{Tritrichomonadea}$),拥有三个向前的鞭毛(极少四个)和波状膜结构。这个家族的杰出代表是胎三毛滴虫($T.~foetus$),可以感染猫咪等宠物导致腹泻,感染牛、猪等动物的生殖道导致流产。这种疾病被称为动物滴虫病,传染性很强且容易复发,是畜牧业棘手的难题。我们甚至已经在人类患者的肺部发现了它的存在,表明可能存在适应人类宿主的寄生虫株(Duboucher et al. 2006)。
(上图:$T.~foetus$ 的细胞结构和显微图像)
另一个主要寄生脊椎动物的类群是双核阿米巴虫纲($\text{Dientamoebea}$),它正是三毛滴虫的姐妹群,只不过从形态上完全看不出二者之间的关系。最常见的脆弱双核阿米巴虫($D.~fragilis$),长相与同门几乎所有物种都大相径庭。顾名思义,它有两个细胞核,通过伪足运动,根本没有鞭毛(被认为是在演化中丢失的),且这似乎是它唯一可活动的形态。若不是副基体出卖了它的身份,它可能到现在还被归类为变形虫:
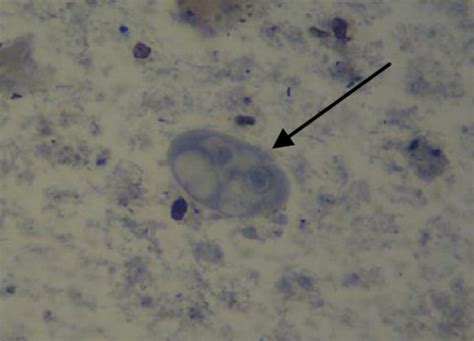
它或许是最成功的人类肠道寄生虫之一,不同地区的感染率可以从 $1\%$ 飙到 $90\%$,许多发达国家的感染率甚至超过发展中国家。作为寄生虫,它最大的优势或许就是“作风优良”,大多数人感染之后都没有症状,就算有症状也都是慢性的,且潜伏期很长,以至于很多时候我们都会忽略它的存在。
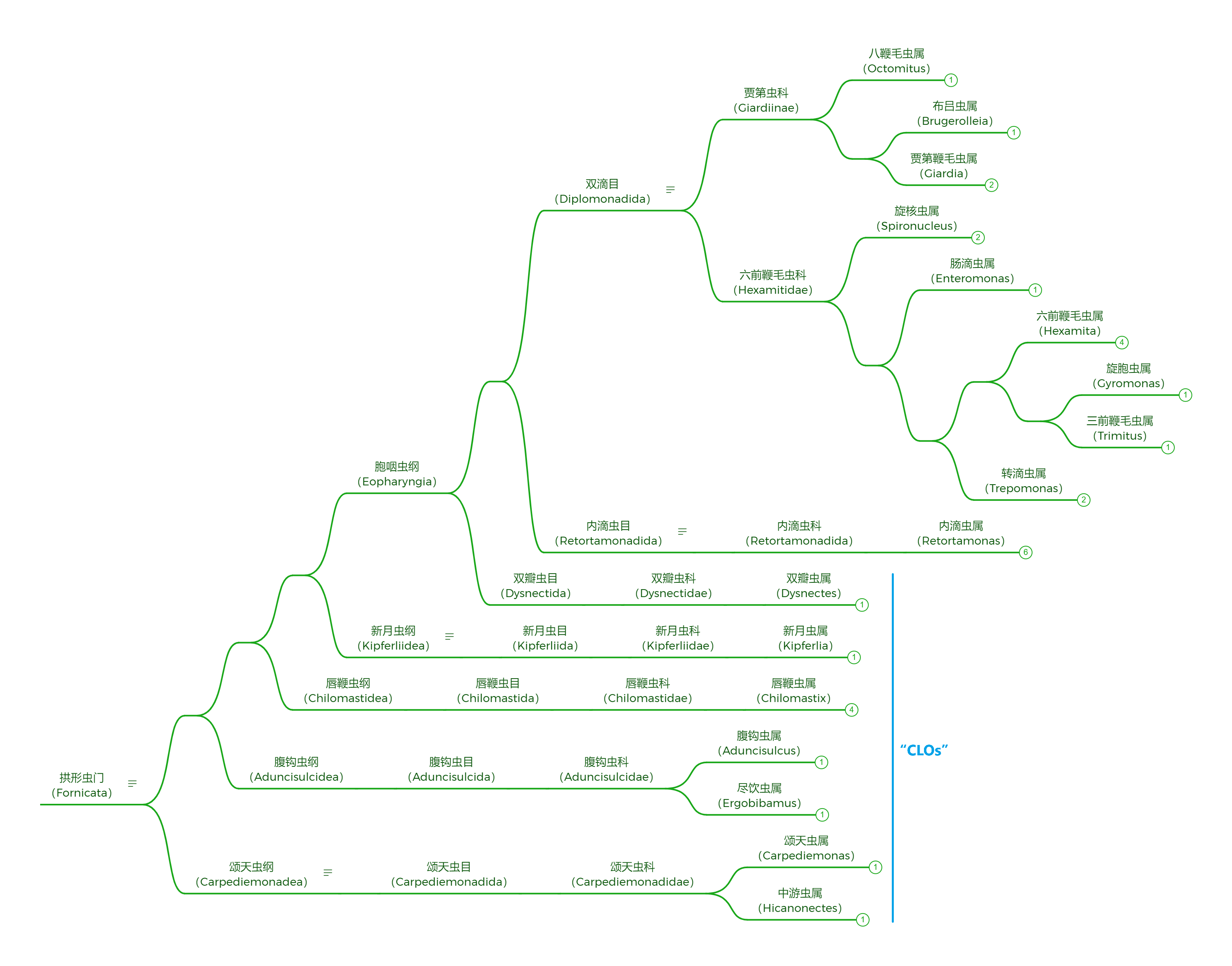
同科不同属的组织单胞滴虫($H. meleagridis$)则具有非常复杂的生命历程。其第一宿主是一些线虫,此时的它非常小,可以直接进入线虫的卵从而感染后代线虫。如果这些卵被第二宿主——蚯蚓吞食,它就会顺势钻入蚯蚓体内。如果最终宿主——鸡形目鸟类吃掉了蚯蚓,或者是经由一些寄生性线虫传播给了鸟类宿主,那它就会变成鞭毛虫形态,进入盲肠组织中大量繁殖。它还会迁移到鸟类的肠黏膜中,变成与 $D.~fragilis$ 类似的变形虫形态(但只有一个细胞核),并造成大面积的组织坏死。
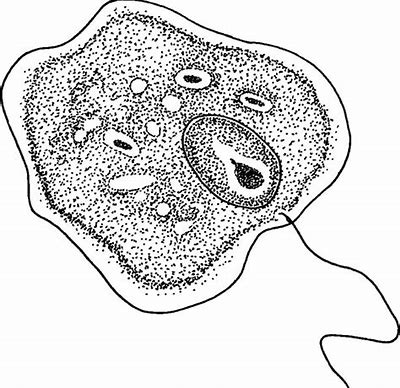
(上图:$H.~meleagridis$ 的鞭毛虫形态手绘。可以看到它只有一根鞭毛,且轴柱非常短,隐藏于体内)
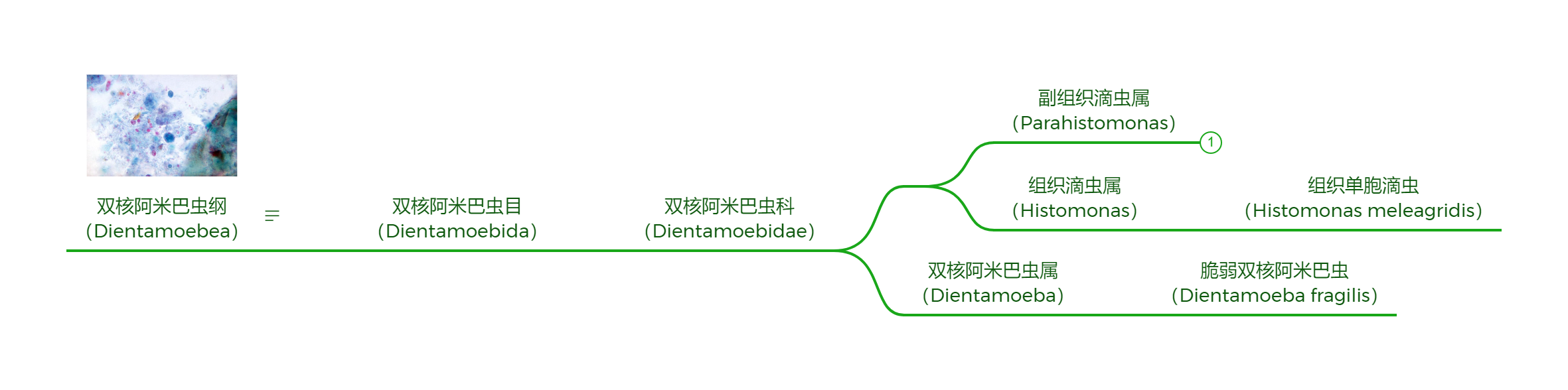
(上图:双核阿米巴虫纲的系统发育树)
对于火鸡等鸟类来说,$H.~meleagridis$ 是极为致命的,一旦感染就有超过 $80\%$ 的死亡率。或许是为了配合自己的致命性,$H.~meleagridis$ 还可以直接通过鸟类之间的性行为传播。如果继续演化下去,说不定它们中的一部分会彻底抛弃线虫和蚯蚓,变成鸟类的专属寄生虫。或许,$D.~fragilis$ 也走过了类似的演化道路,并在成为人类的专属寄生虫之后抛弃了其他形态,从而变成了今天这副样子。
一个佐证是,$D.~fragilis$ 经常与蛲虫($E.~vermicularis$,一种同样寄生于人类肠道中的线虫)一起被发现于病人体内,这可能说明 $D.~fragilis$ 仍然有通过线虫传播的能力。有人进一步提出,双核阿米巴虫纲的祖先可能就是寄生于线虫体内的,后来通过一些寄生性线虫传播给了脊椎动物。在后续的演化中,它们中的一些掌握了不依靠线虫的传播方法,便逐渐抛弃了线虫而专攻脊椎动物,进而变成了今天的模样。
$\text{Cadamassta}$ 总纲中另外两个关系密切的类群是螺旋滴虫纲($\text{Spirotrichonymphea}$)和冠滴虫纲($\text{Cristamonadea}$)。它们都生活在白蚁的肠道内(它们的共同祖先应当也是),到底是寄生还是共生不太好说,毕竟二者没有明显边界。螺旋滴虫纲内物种形态类似,都有很多鞭毛,排列为右手螺旋状:
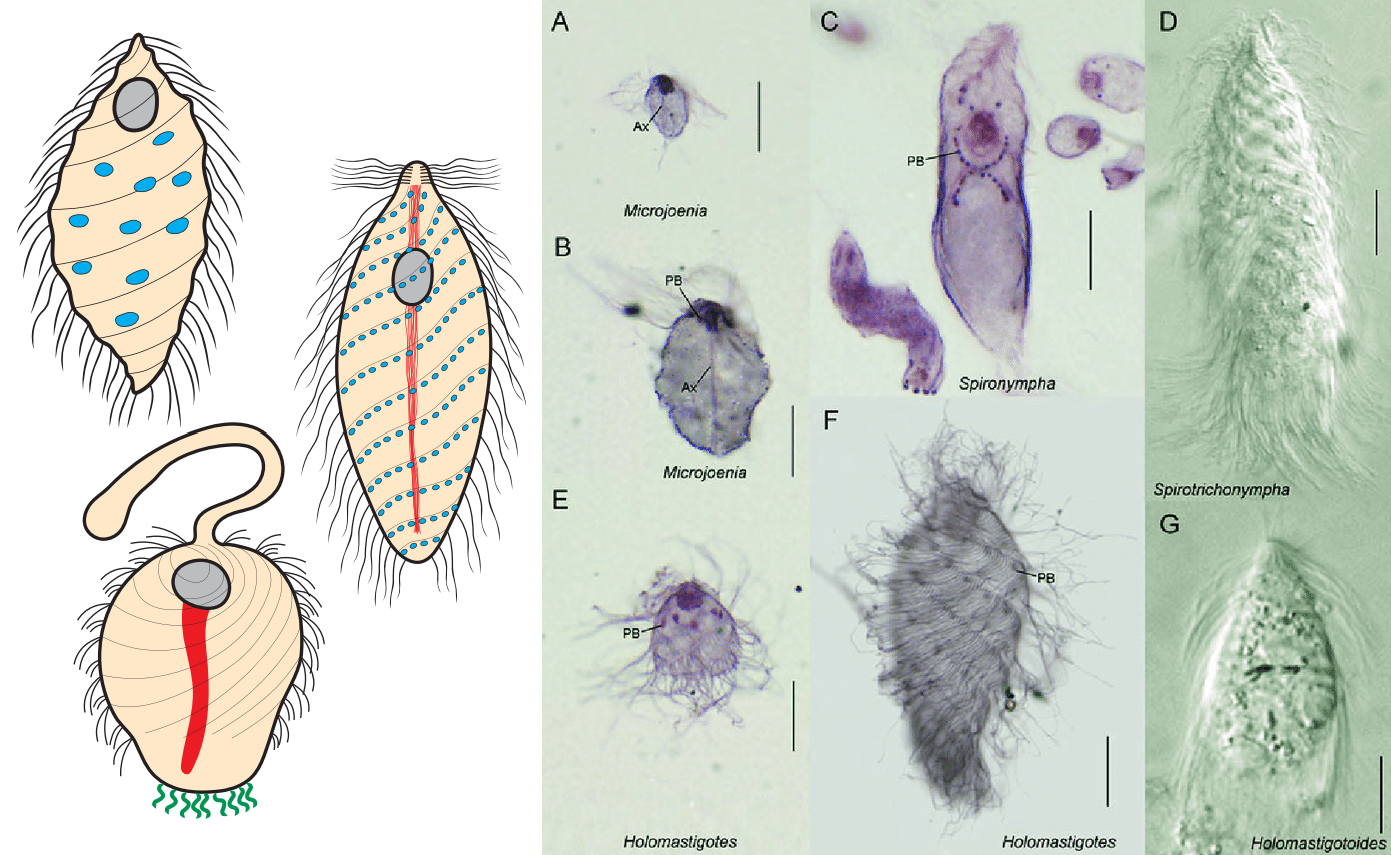
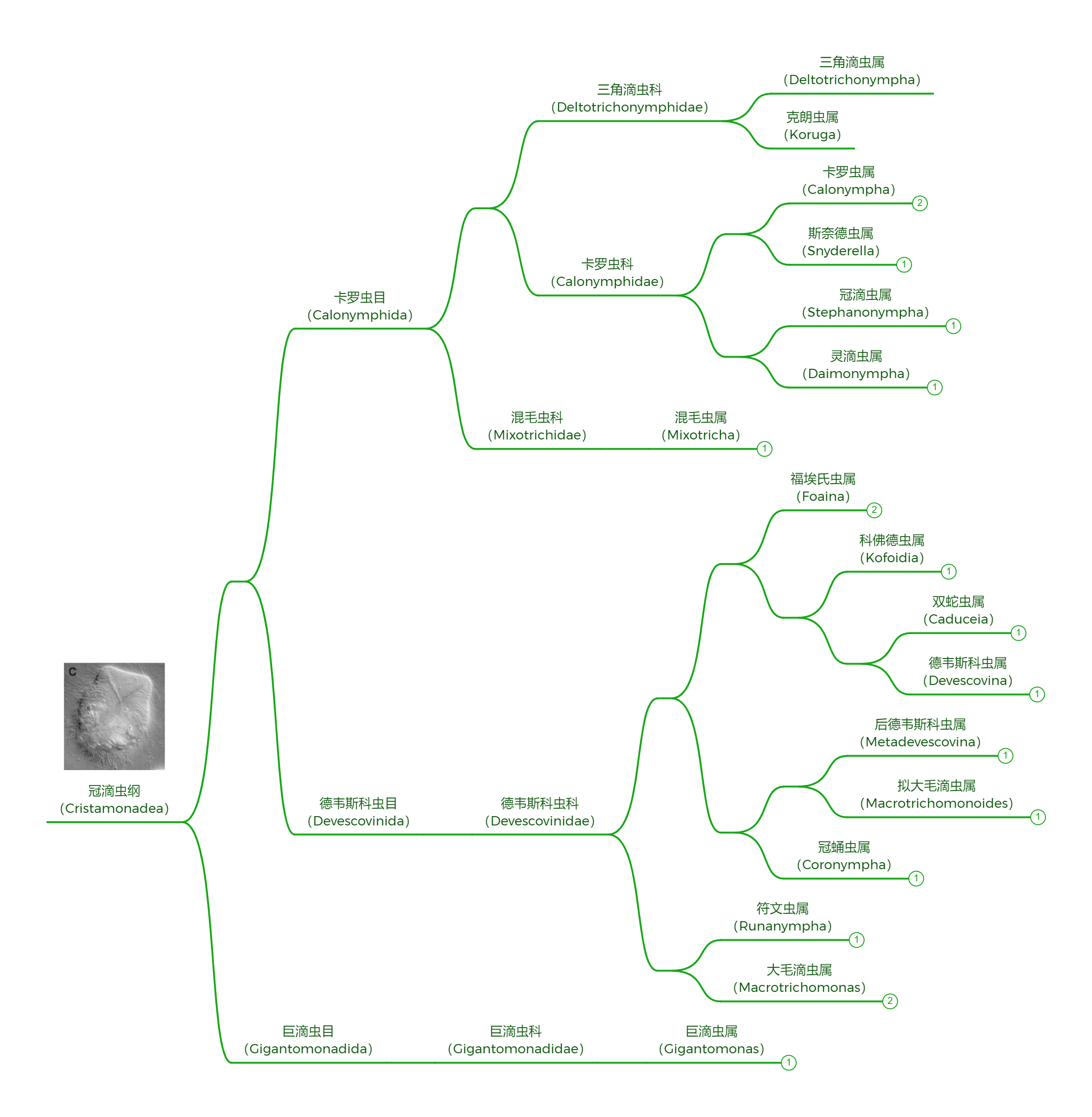
冠滴虫纲的形态就更多样一些。纲基部的巨滴虫目($\text{Gigantomonadida}$)只有四根鞭毛,三条向前,一条明显更粗的鞭毛向后;德韦斯科虫目的大部分成员也只有四根鞭毛,但是有形态完全不同的例外:符文虫属($Runanympha$)。它有超多的鞭毛,但是都集中在顶部,而不是像螺旋滴虫一样覆盖整个细胞。
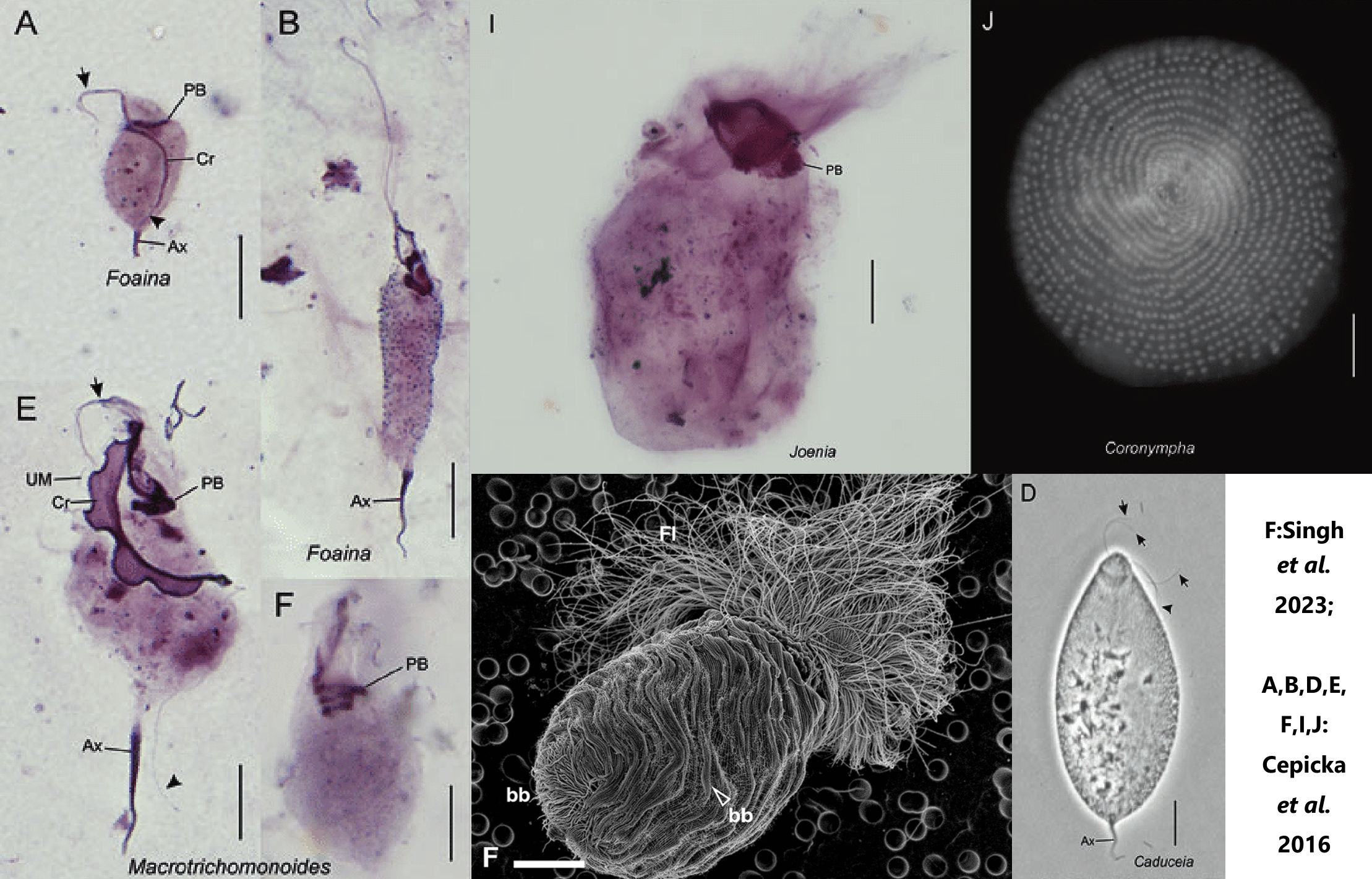
(上图:德韦斯科虫目部分成员。黑背景 $\text{F}$ 图为符文虫属)
当然,冠滴虫纲内最重量级(字面意思)的类群当属卡罗虫目,因为它们的通常形态为合胞体,体型自然也比纲内其他类群大。在我看来,其中最有意思的当属灵滴虫属($Daimonympha$)。它们的细胞前端可以快速旋转,其中的细胞核也跟着一起转,如下面的动图:

(上动图:$D.~friedkini$)
我们尚不知道这种旋转是如何实现的,也不知道在此过程中细胞膜如何进行剪切、细胞骨架如何变化,更不知道这种旋转是如何演化而来,它对于生存究竟有什么意义。事实上,我们对于这些类群的了解都极为有限。
在系统发育树上,冠滴虫纲有几十个属的位置是未知的。其他已被确认的类群之间的关系如下图所示:
总的来说,$\text{Cadamassta}$ 总纲的演化解释起来是比较简单的,它们很可能是由同一个内生的祖先演化而来。然而在副基体门的另一大分支中,情况就变得复杂了起来。
处于这一分支(并没有被正式命名,个人觉得可以叫毛滴虫总纲)基部的是次毛滴虫纲($\text{Hypotrichomonadea}$)。它光看外表很难和毛滴虫区分,但细细观之,你会发现它没有被称为下基体($\text{infrakinetosomal body}$)的细胞器,而毛滴虫纲所有物种都有下基体。
(上图:$H.~acosta$)
次毛滴虫纲现存的所有物种都共生于动物肠道中,结合其在演化树上的位置,难道毛滴虫总纲的内生也是单起源的吗?或者说,难道副基体门的唯一一次向内生的转变是在宾巴维虫分化之后,两大总纲分化之前?
事实并不是这样。$2022$ 年的一项研究在毛滴虫纲($\text{Trichomonadea}$)内部发现了许多自由生活的物种,而系统发育告诉我们,它们相比于内生的毛滴虫更加靠近基部,这在下图中可以看得很清楚:
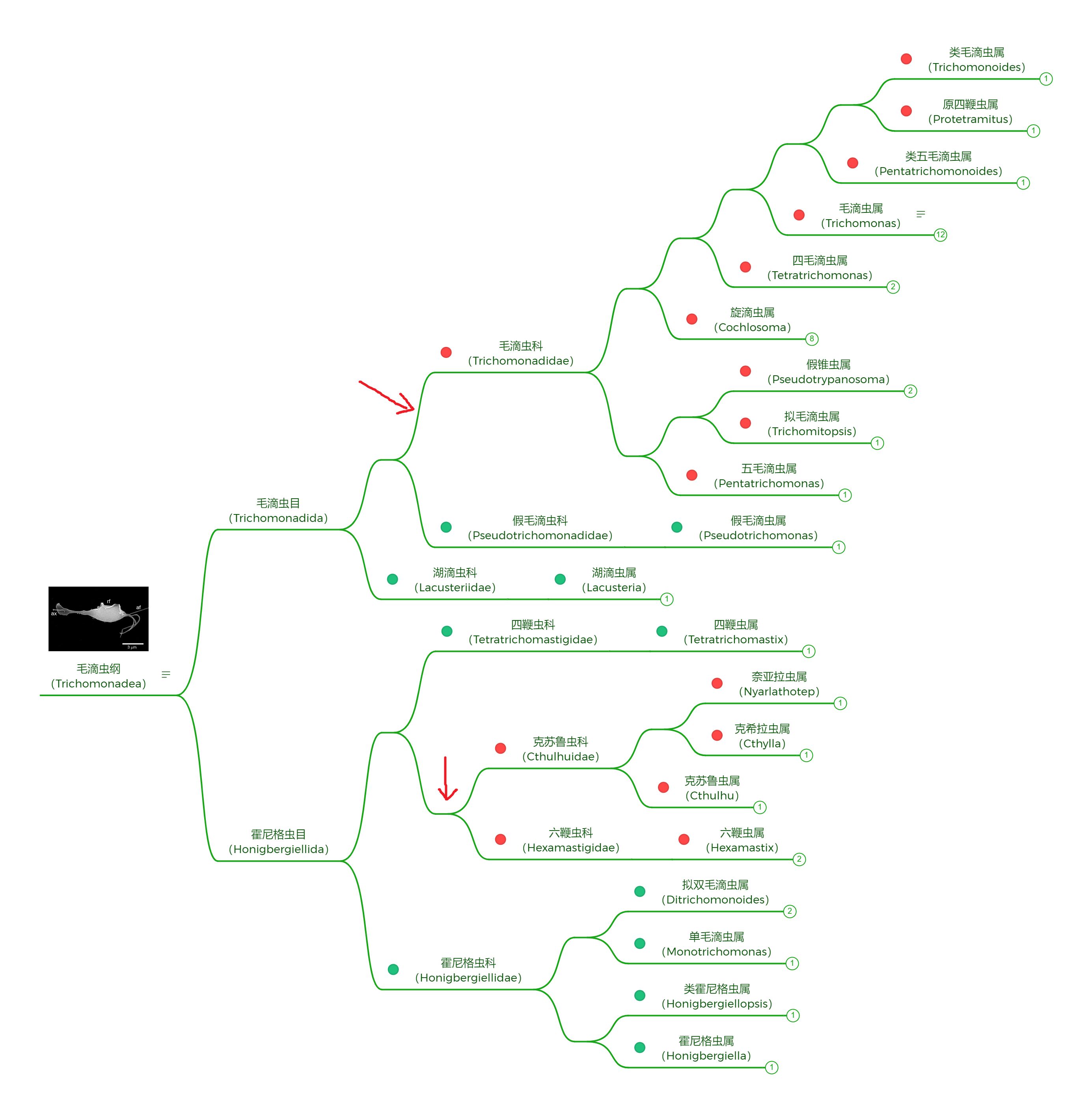
(上图中:绿色标记表示自由生活,红色标记表示内生)
由此推测,毛滴虫纲的祖先是自由生活的,而在后续演化中出现了两次独立的向内生的转变。在形态上,自由生活和内生的毛滴虫没有太大区别。单毛滴虫属($\text{Monotrichomonas}$)比较有特点,它只有一个向前的鞭毛:
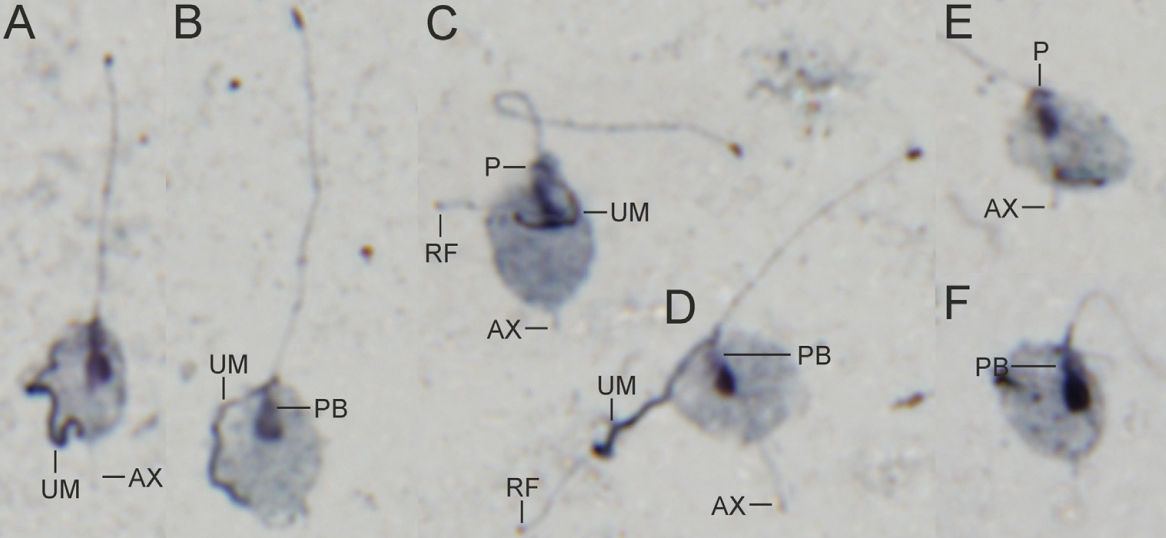
(上图:$M.~transatlantica$)
假毛滴虫科($\text{Pseudotrichomonas}$)与毛滴虫科是姐妹群,两者在形态上也更加相近。$P.~buchari$ 有四条几乎等长的前鞭毛,另有复鞭毛形成波状膜:
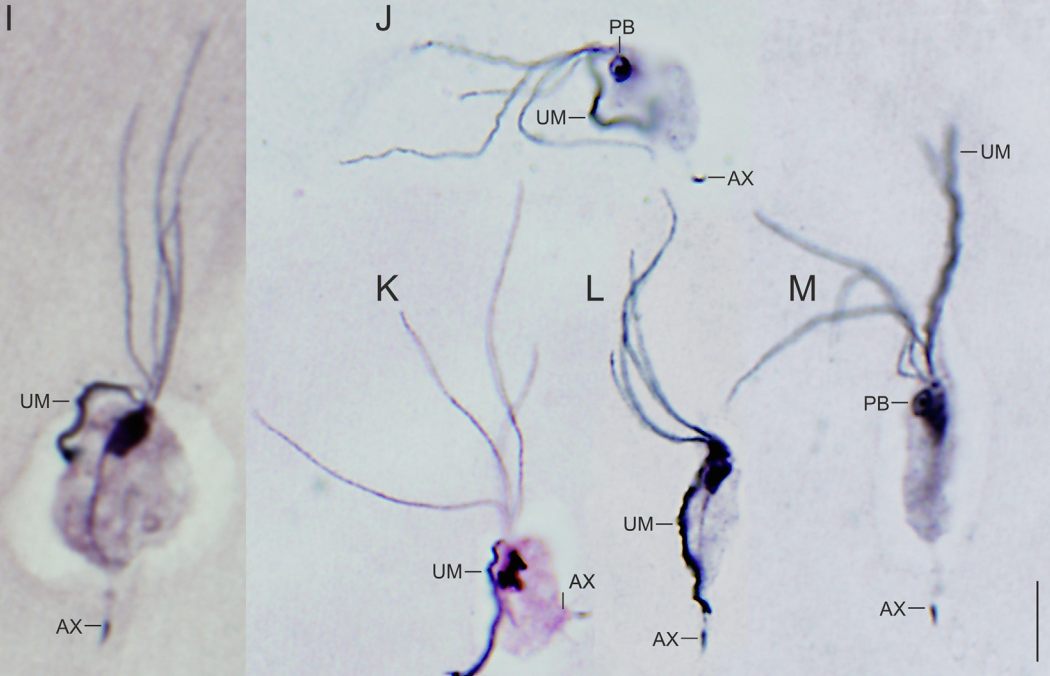
在内生毛滴虫中,克苏鲁虫科($\text{Cthulhuidae}$)是最近被发现的,虽然我怎么也想不通它们和克苏鲁到底有什么关系。其中的克苏鲁虫属($\text{Cthulhu}$)共生于哥伦比亚的一种低等白蚁体内帮助其消化纤维素,它将自己的十几根鞭毛捆成一到两束,并以一种难以描述的古怪姿势运动:
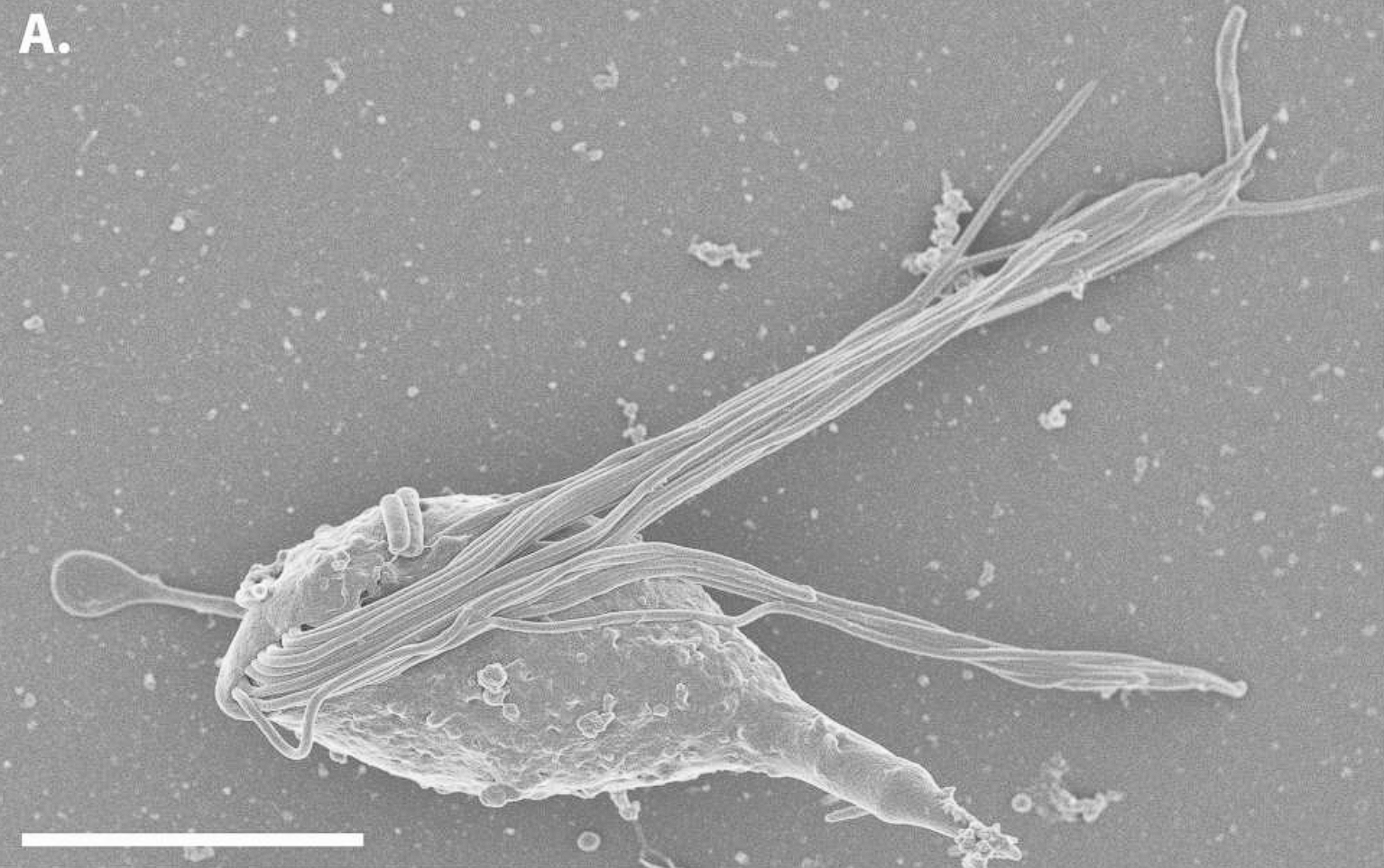
(上图:$C.~macrofasciculumque$)
当然,以上这些类群在毛滴虫科($\text{Trichomonadidae}$)面前,都只能算是一伙杂鱼。它们中的很多都是寄生虫,主要寄生脊椎动物。其中最知名、对人类威胁最大的当属阴道毛滴虫($T.~vaginalis$)。它通过性行为传播,并造成滴虫性阴道炎。据估计,世界范围内每年感染 $T.~vaginalis$ 的人超过一亿($2011$ 年的研究,现在应该少一些了吧),但大多数感染者都没有症状。今天的我们可以使用甲硝唑等药物干掉这玩意,而在古代,虽然没有详细记录,但也可以推测它是很多新生儿死亡的元凶。
形态上,$T.~vaginalis$ 长得像插了根尾巴的纺锤,轴柱突出,有四根前鞭毛,另有复鞭毛形成波状膜,是经典的毛滴虫结构:
除了毛滴虫之外,近年还发现冠状单胞虫纲($\text{Lophomonadea}$)的 $L.~blattarum$ 也可以成为人类的病原体。它的细胞呈现出卵形或叶形,顶部有一个像花萼一样的环状结构,几簇鞭毛附着于其上:
自然状态下,$L.~blattarum$ 共生于蟑螂的肠道内,但我们在一些肺炎患者体内也发现了它。这非常奇怪,因为我们仍无法解释它是怎么进入人体的。它引起的病症也不严重,所以貌似也没什么人关注。
与冠状单胞虫纲关系最近的就是著名的披发虫纲($\text{Trichonymphea}$),二者可能来源于同一个内生祖先。事实上,在此前的分类里,冠状单胞虫披发虫纲的一个目,也不知道为什么要独立出来。只要见过它们的样子,你就知道“批发”这一译名是多么形象。它们总是拥有很多鞭毛,两侧对称分布,大部分物种的鞭毛更是遍布全身(看起来还有点可爱?):
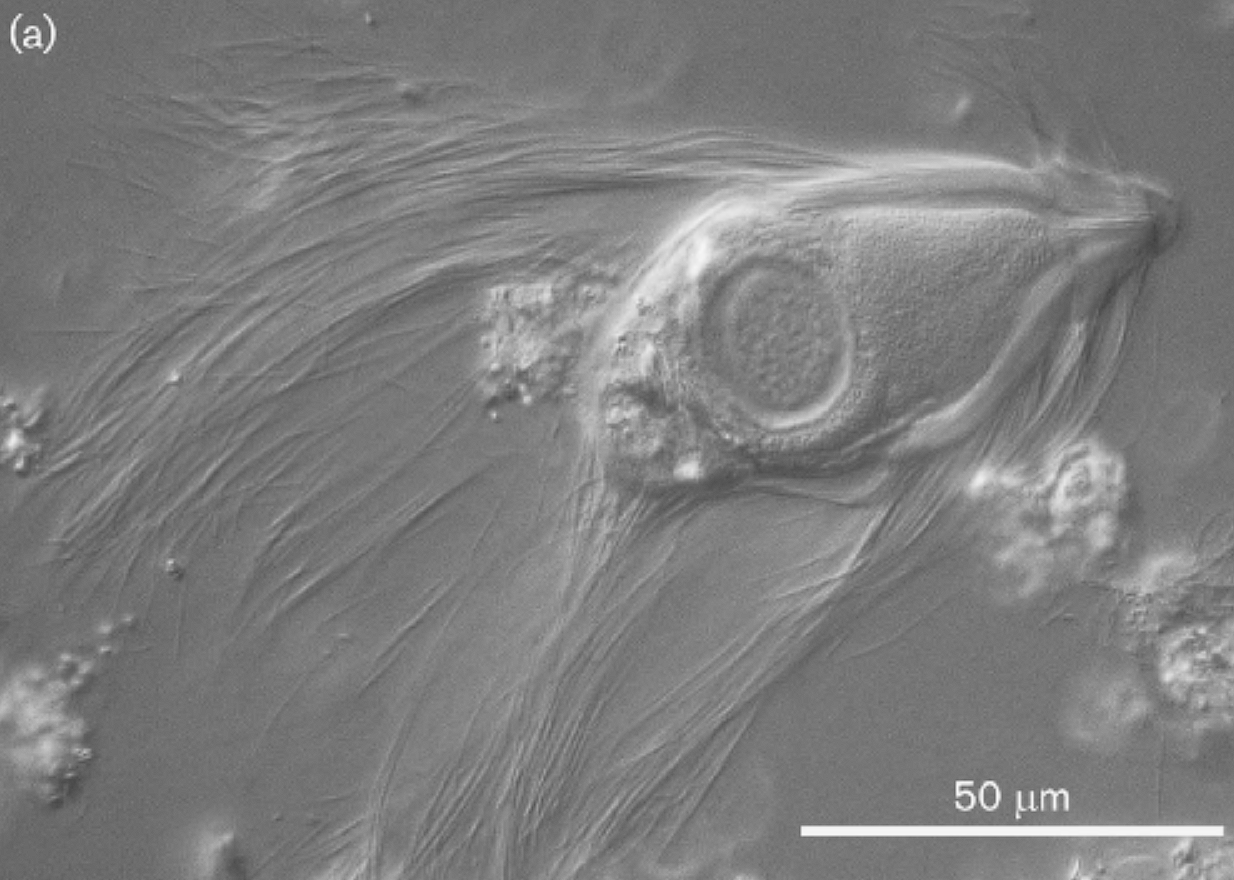
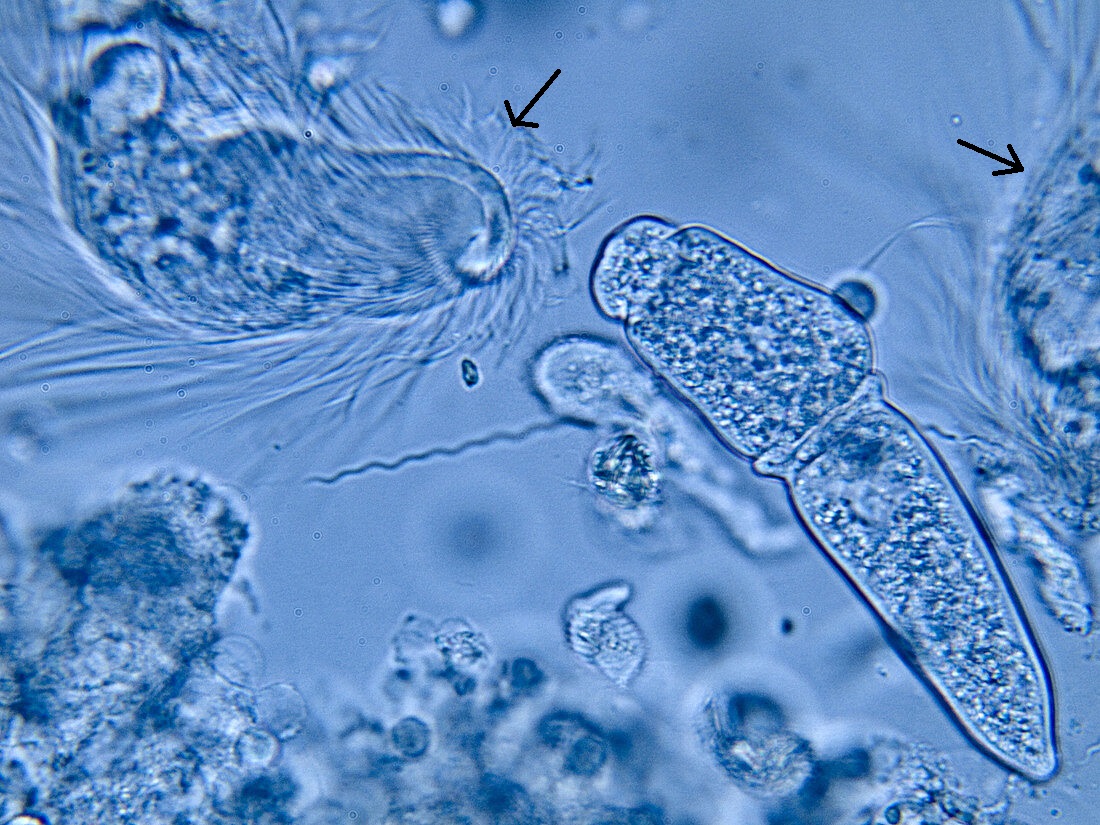
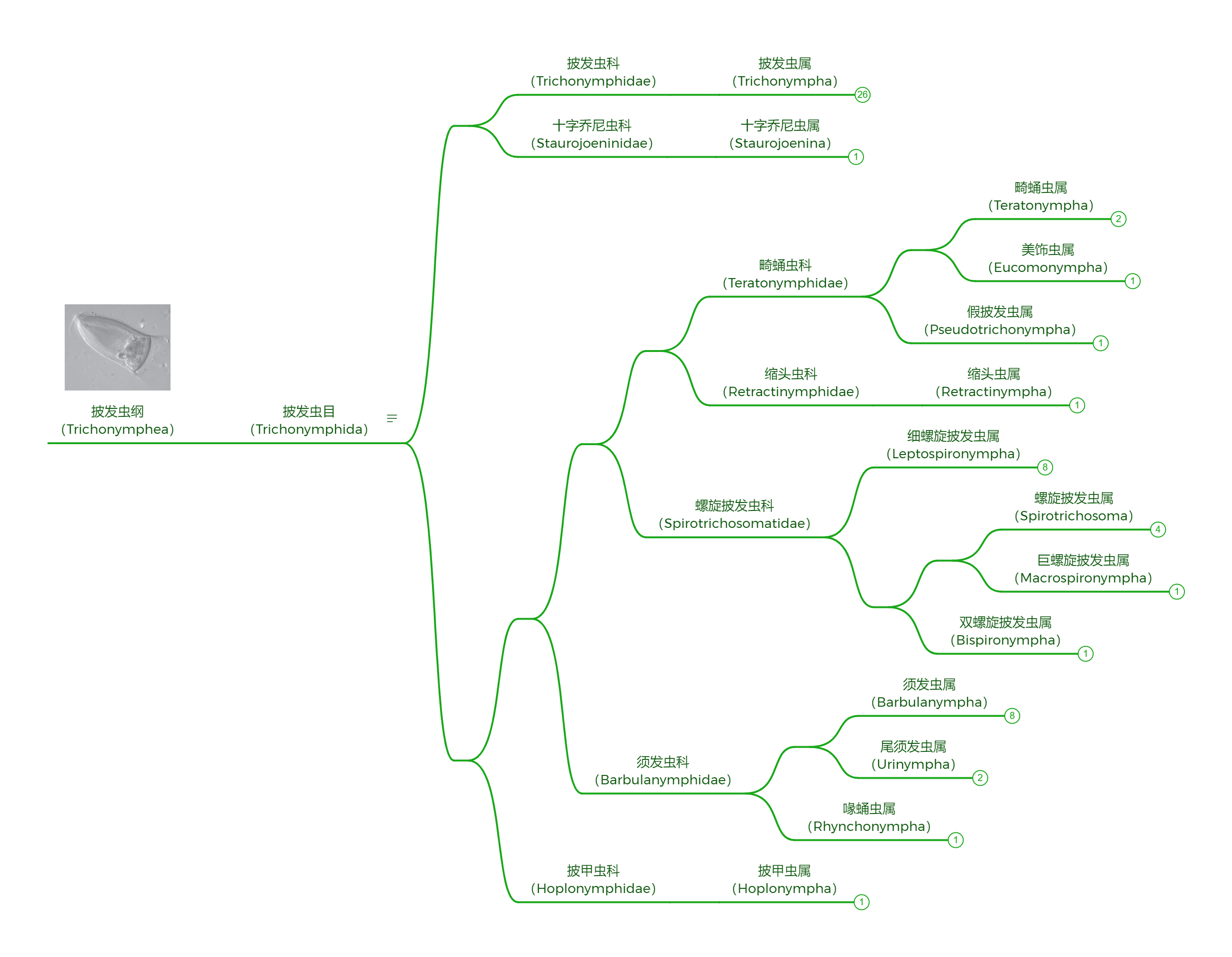
作为最出名的昆虫肠道共生者之一,它们可以为食木昆虫提供消化纤维素的酶。依靠这种本领,它们顺利地扎根在了白蚁、隐尾蠊等蜚蠊目昆虫的肠道内,进而演化出了一个庞杂的家族。下面的系统发育树展示了其中被详细研究的十四个属:
甚至于,披发虫还有已被确认的化石物种。缅甸古披发虫($Burmanymphus$)于 $2009$ 年被发现于一块来自早白垩世的琥珀中的蟑螂的肠道内,在形态上虽然和其他披发虫类群都有些区别(所以被单立为一科),但也大差不差:
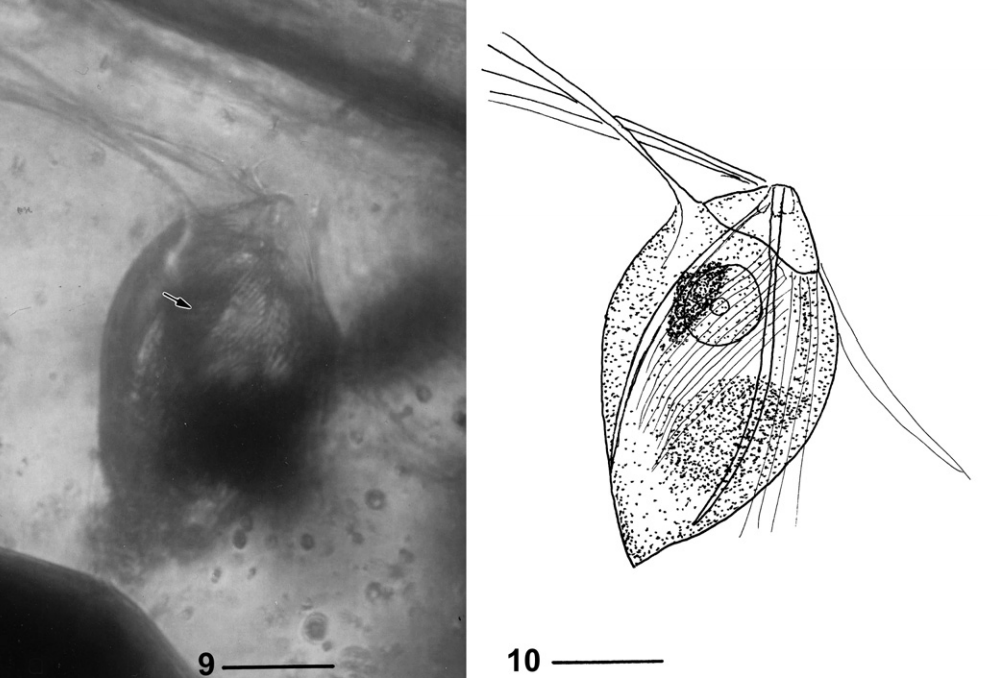
(上图:$B.~cretacea$)
同一化石中还发现了冠滴虫纲德韦斯科虫属($Devescovina$)和前轴柱门锐滴虫属($Oxymonas$)的物种,以及一个形态奇异、根本不知道归类到哪里的物种,会在后文讨论。
奈何,昆虫们也不是总是愿意被中间商赚差价的。白蚁科($\text{Termitidae}$),也就是高等的、狭义的白蚁演化出了自己的纤维素酶,从而将披发虫赶了出去。披发虫自己也有独特的内共生细菌(也有一些是体外共生),其中一种细菌很可能拥有固氮的能力,从而为自己的宿主,以及宿主的宿主提供食物中缺乏的氮元素;另一种细菌很可能来自脱硫弧菌属($Desulfovibrio$),为披发虫提供了一整套硫酸盐代谢通路。从演化上来看,披发虫和这些细菌的共生关系很可能在它们的共同祖先那儿就确立了。或许就是凭借着这一手黑科技,披发虫成为了后滴虫中最为成功的昆虫肠内共生者。
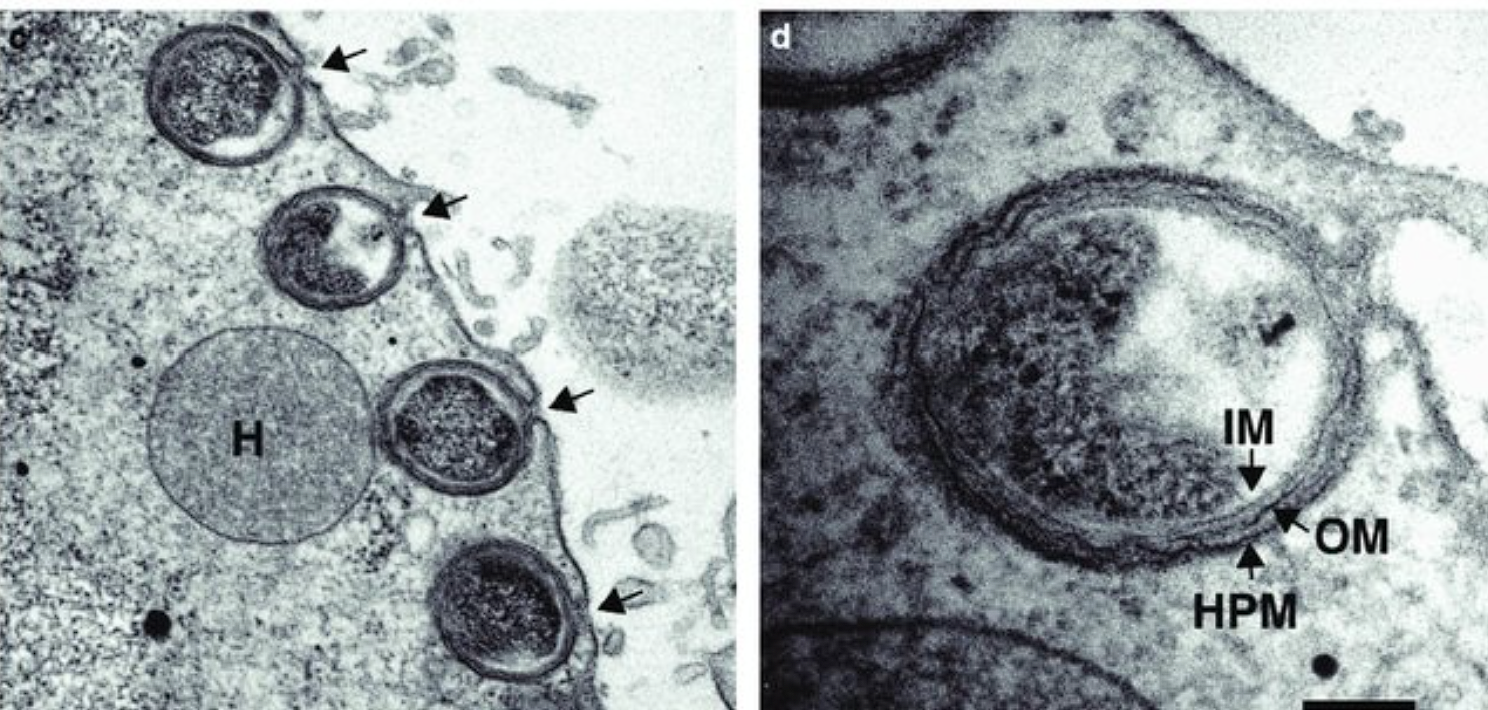
(上图:与披发虫共生的细菌 $D.~trichonymphae$)
到这里,我们已经大致完成了关于副基体门的讨论,它们与厌氧变形虫门共同构成了后滴界的一个至高分支:$\text{AP}$ 类群;另一个至高分支则被称为 $\text{FBP}$ 类群,也是其下三门首字母的缩写。
位于 $\text{FBP}$ 类群最基部的则是前轴柱门($\text{Preaxostyla}$)。既然提到了,就必须要吐槽一下目前该类群的正式名 $\text{Anaeromonada}$,翻译过来叫厌氧滴虫门。可是,所有后滴虫都是厌氧的,这个名字就仿佛没事找事一般,遇到眼力不行的(比如我)还容易和厌氧变形虫混淆。最关键的是,它没有突出该类群的形态特征。
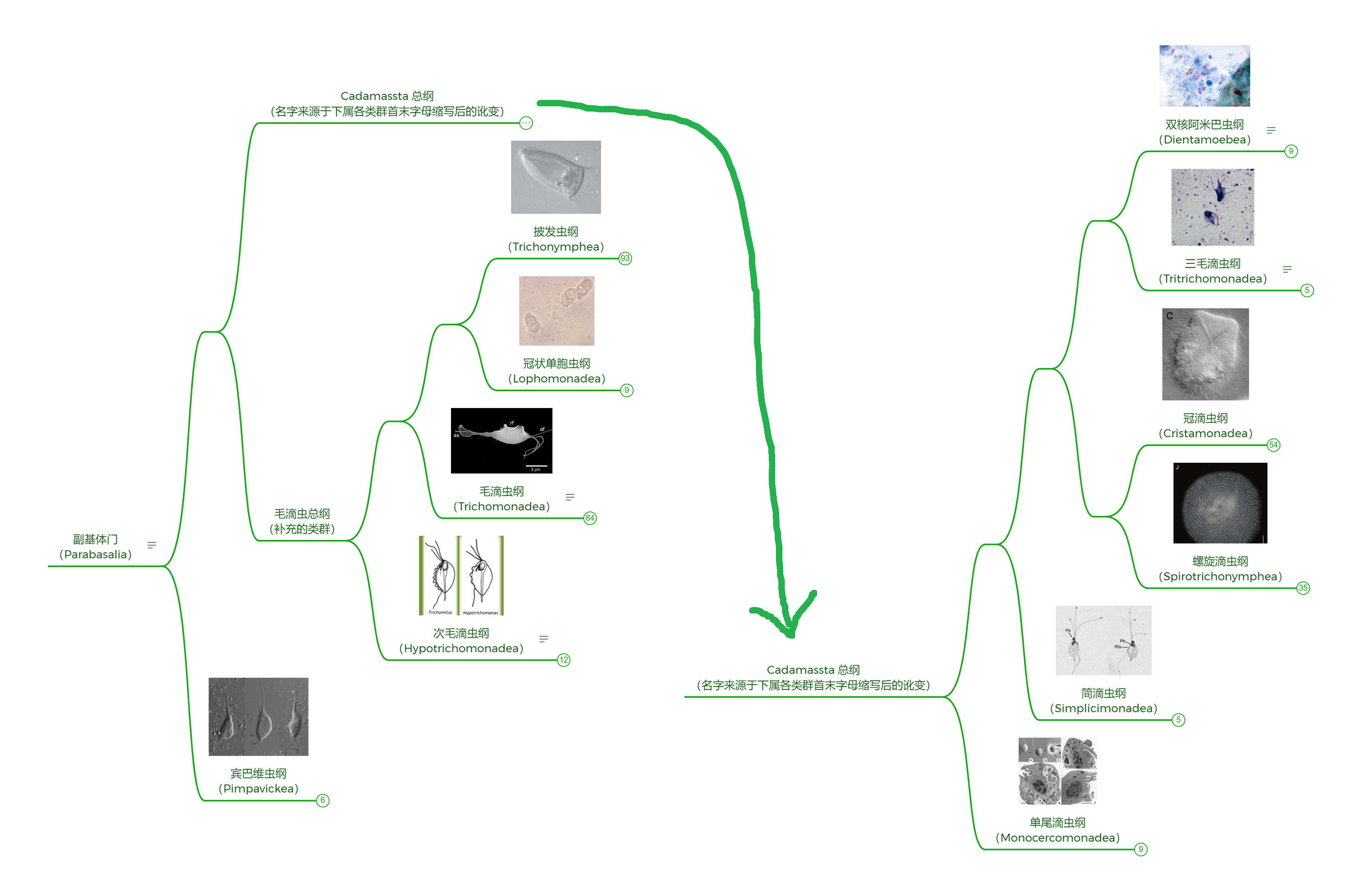
我们之前在描述形态时提到过“轴柱”($\text{axostyle}$)一词,这是一种由维管形成的束状结构,可能起到辅助运动的作用。副基体门和前轴柱门都有轴柱,但很可能不是同源的。最明显的区别,前轴柱门的轴柱在细胞前端切分鞭毛基部,而副基体门很多类群的轴柱向后突出;后来的研究又指出,二者的轴柱中维管的组装方式截然不同,为这一观点提供了强有力的支持。
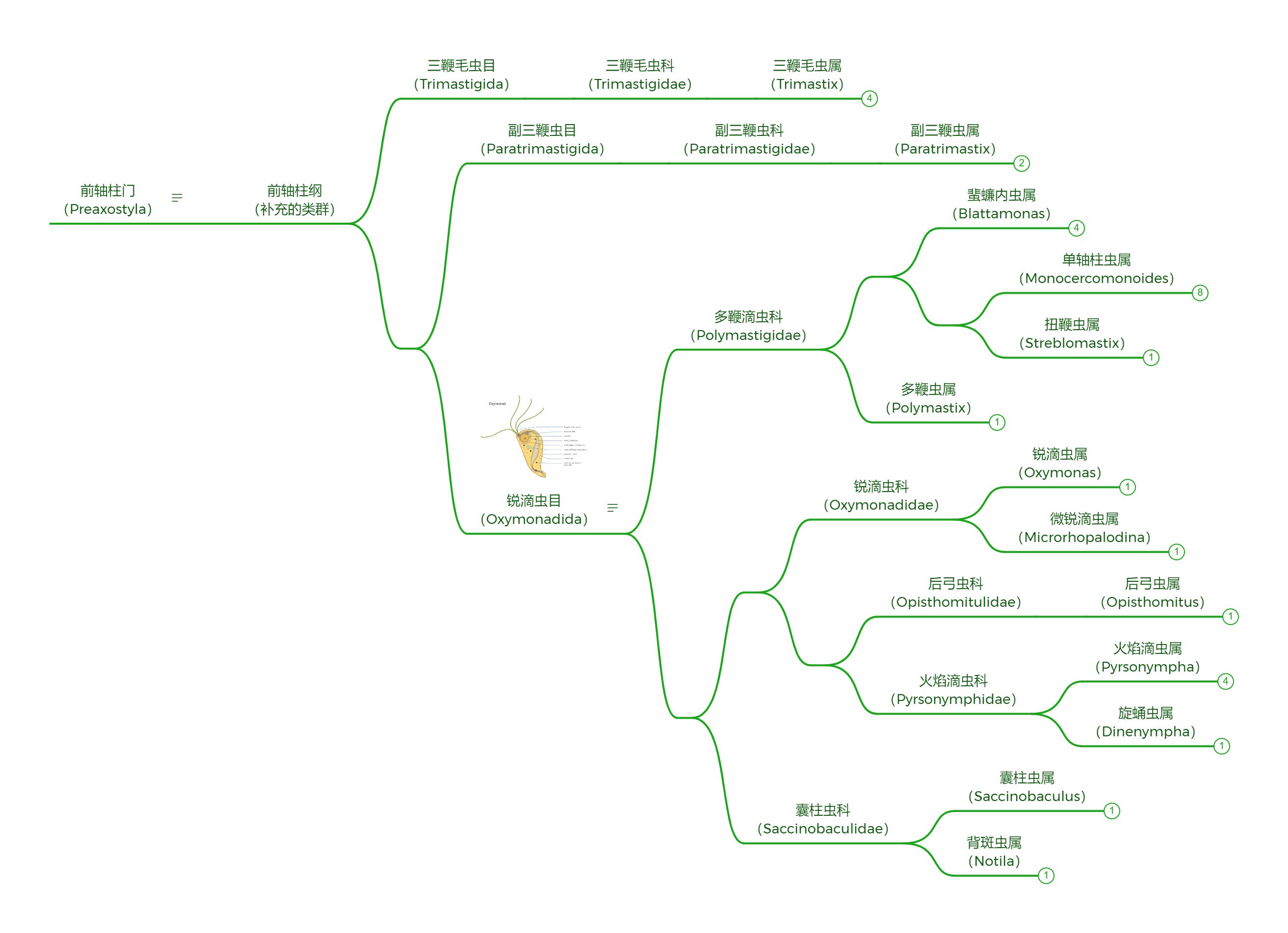
前轴柱门内类群相对较少,$2015$ 年的系统发育研究将其划分为三个目,如下图所示:
处于基部的三鞭虫属($Trimastix$)和副三鞭虫属($Paratrimastix$)是自由生活的。它们有四根鞭毛,三条向前,一条向后,还有一个用于辅助进食的摄食槽(其实许多古虫都有)。前者生活于海洋中,而后者分布于淡水,二者都以原核生物或腐烂的尸体为食。

(上动图:三鞭虫属的 $T.~marina$ 活体珍贵片段)
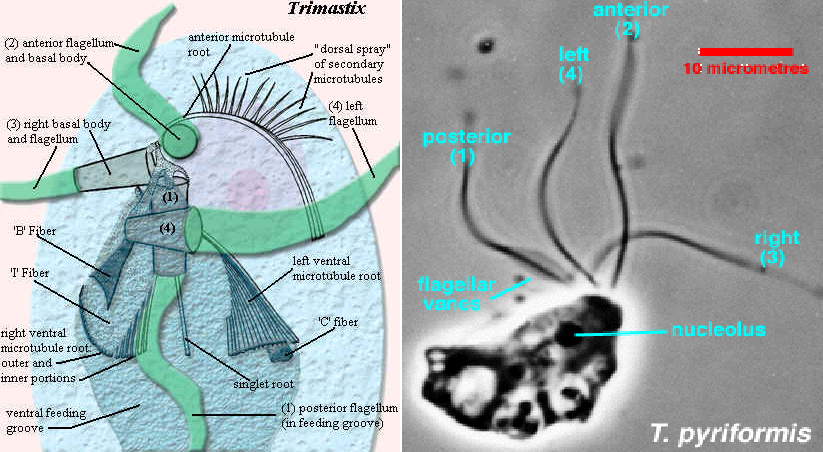
(上图:$T.~pyriformis$,现属副三鞭虫属,更名为 $P.~pyriformis$)
由于形态上几乎没有区别,所以能把副三鞭虫独立出来还是分子生物学的魅力时刻。前轴柱门中,绝大部分物种都在锐滴虫目($\text{Oxymonadida}$)。它们或许是与披发虫趋同演化,同样生活在低等白蚁等食木昆虫的消化道中,帮助它们消化纤维素。形态上,锐滴虫退化了摄食槽,或许内生的它们也不再需要了。
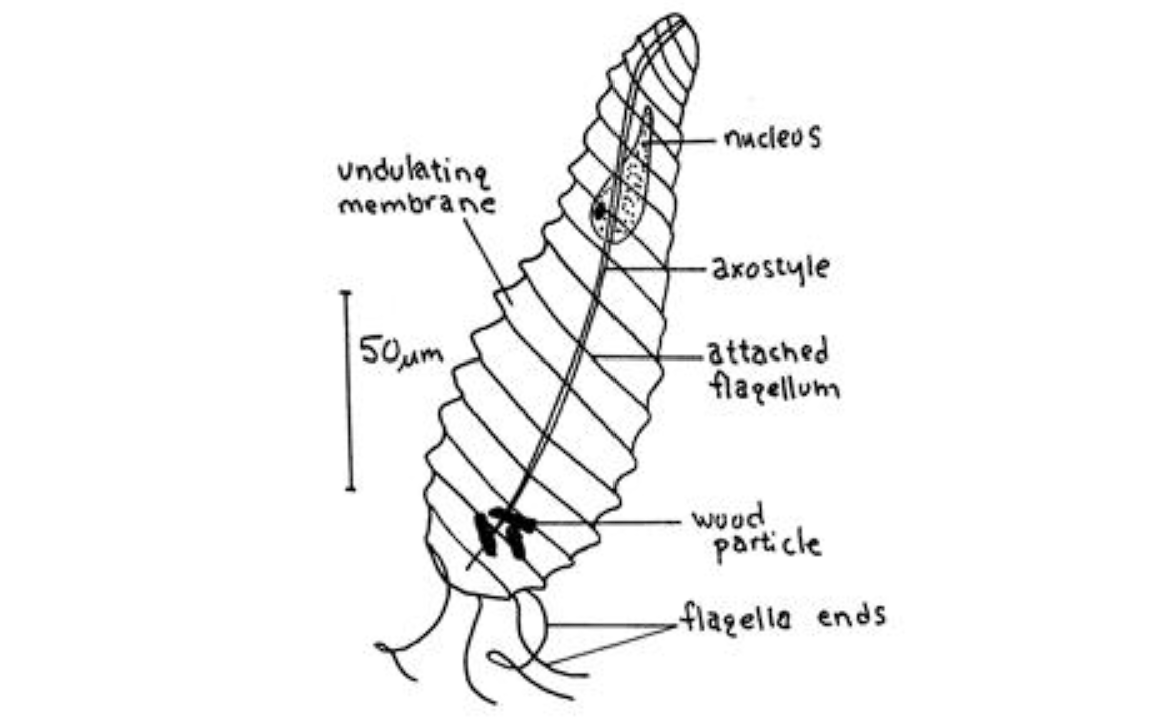
(上图:早期生物学家发现 $O.~granulosa$ 时留下的珍贵手稿)
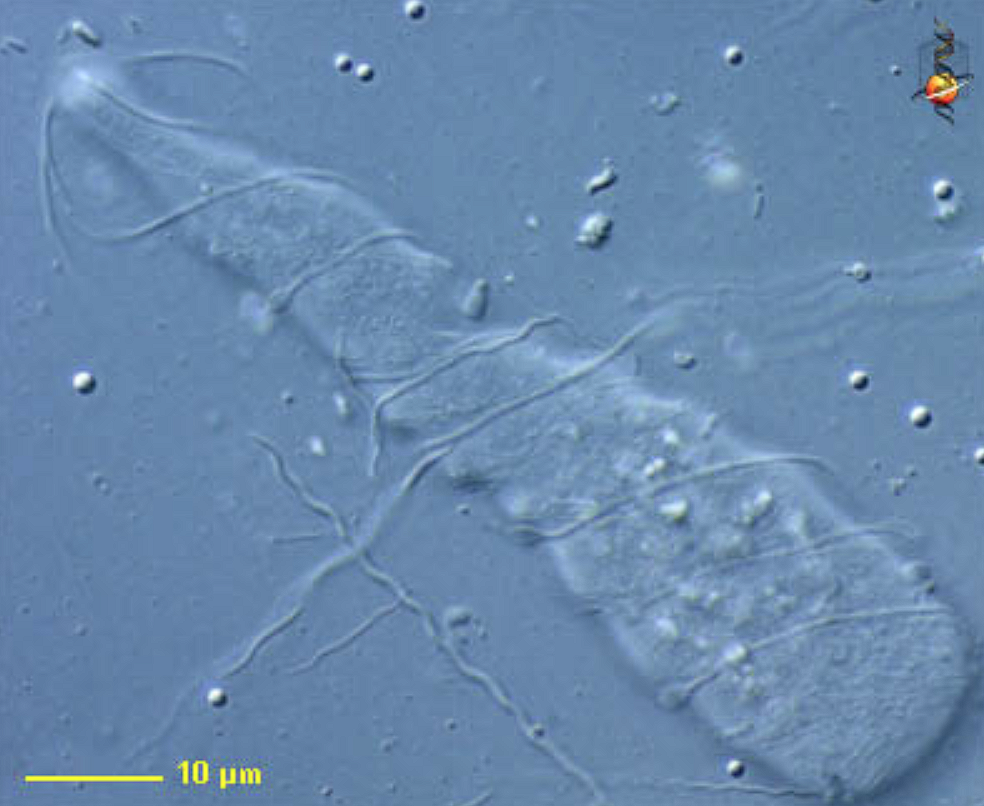
(上图:火焰滴虫属的 $P.~vertens$(到底是谁觉得这玩意长得像火焰啊?))
前轴柱门的演化历程比较简明(因为类群不多)。一开始,它们生活在海洋中,就如最基部的三鞭虫那样;然后,它们中的一部分迁徙到了淡水,以同样的食物为生,今天的副三鞭虫便始终保持着这祖宗之法;而来到淡水,也意味着有更多可能与陆地动物相遇。最终,锐滴虫定居在了蜚蠊目昆虫祖先的肠道内(或许比披发虫更早),与之一起开疆拓土的同时也随之一起演化,最终形成了一个庞杂的家族。今天,门中自由生活的演化支已经几乎凋零殆尽,而锐滴虫自然就接过了前轴柱门正统的大旗。这一假说也与它们有氧呼吸的退化吻合。
但是,一个新发现表明,锐滴虫似乎是有点极端了。对单轴柱虫属($Monocercomonoides$)内物种 $M.~exilis$ 的深入研究表明,它没有任何已知的和线粒体相关的蛋白质,也完全测不出和线粒体相关的基因组。自然地,我们完全观察不到它有任何有氧呼吸的迹象。以后如果有人说,真核生物必定会有氧呼吸,至少得会一点,那你就可以拿这个其貌不扬的小家伙反驳他:
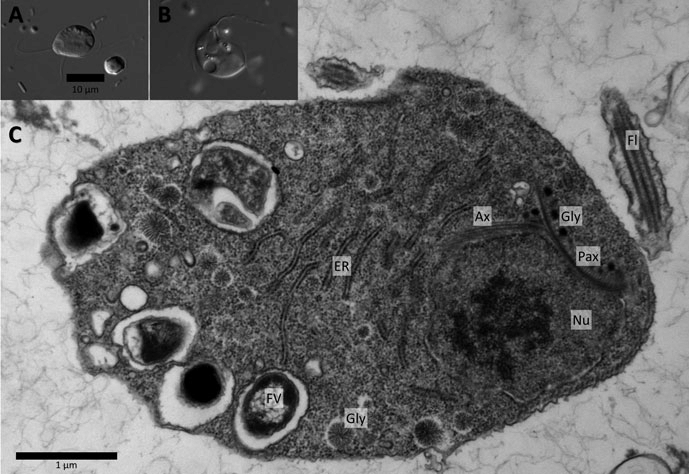
(上图:$M.~exilis$ 的活体图像($\text{A,B}$)与内部构造($\text{C}$))
一些人据此认为,如果真的是线粒体退化造就的,那这种从基因层面上的“清除”还是过于极端,且没有必要。但我坚持认为,这显然不足以驳倒传统观点,毕竟从三鞭虫,到副三鞭虫,再到锐滴虫,与线粒体相关的基因数是递减,这明显是在导向退化的结论。
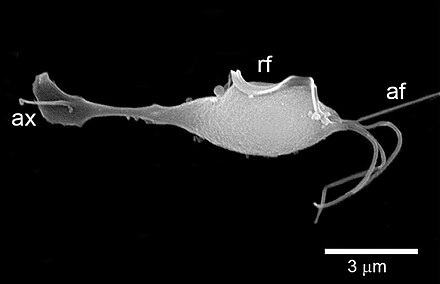
除了前轴柱门外,$\text{FBP}$ 分支内还有两门,其中 $\text{BaSk}$ 门是最近才建立起来的(以后应该会被分配正式名,但总之我写的这一刻还没有)。其下两个类群中,巴塞尔虫($Barthelona$)于 $2000$ 年被发现。它有两个鞭毛,会游泳,自由生活于水体的缺氧区域中:
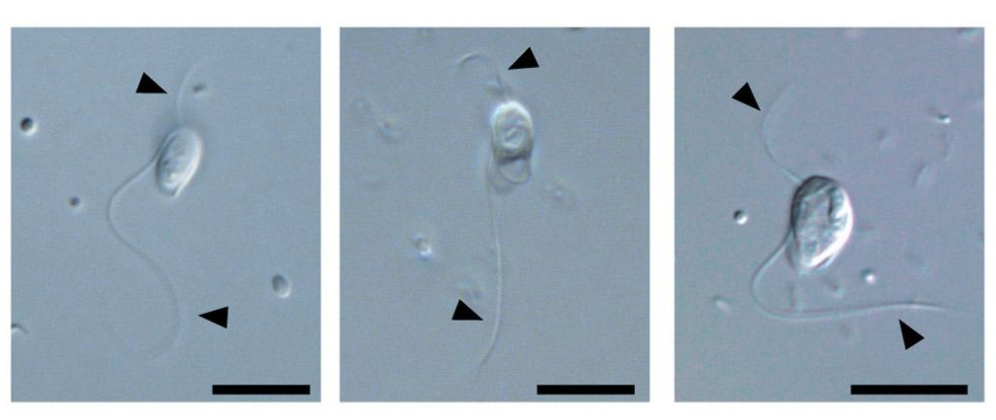
(上图:$B.~vulgaris$)
据说在静止时,它会“以柔和的节奏不停敲打自己的鞭毛”,但可惜没找到视频。门内另一个类群,驼背虫($Skoliomonas$),则在去年才被发现于盐湖和碱湖中,以同样生活在这种极端环境中的原核生物(比如盐杆菌)为食。它的形态比较奇怪,背上像是长了几个瘤子(其实是食物泡,下图的“$\text{fv}$”),有一前一后两个鞭毛,后部那个尖尖的不是突出的轴柱,而是尖刺($\text{spike}$,下图的“$\text{s}$”):
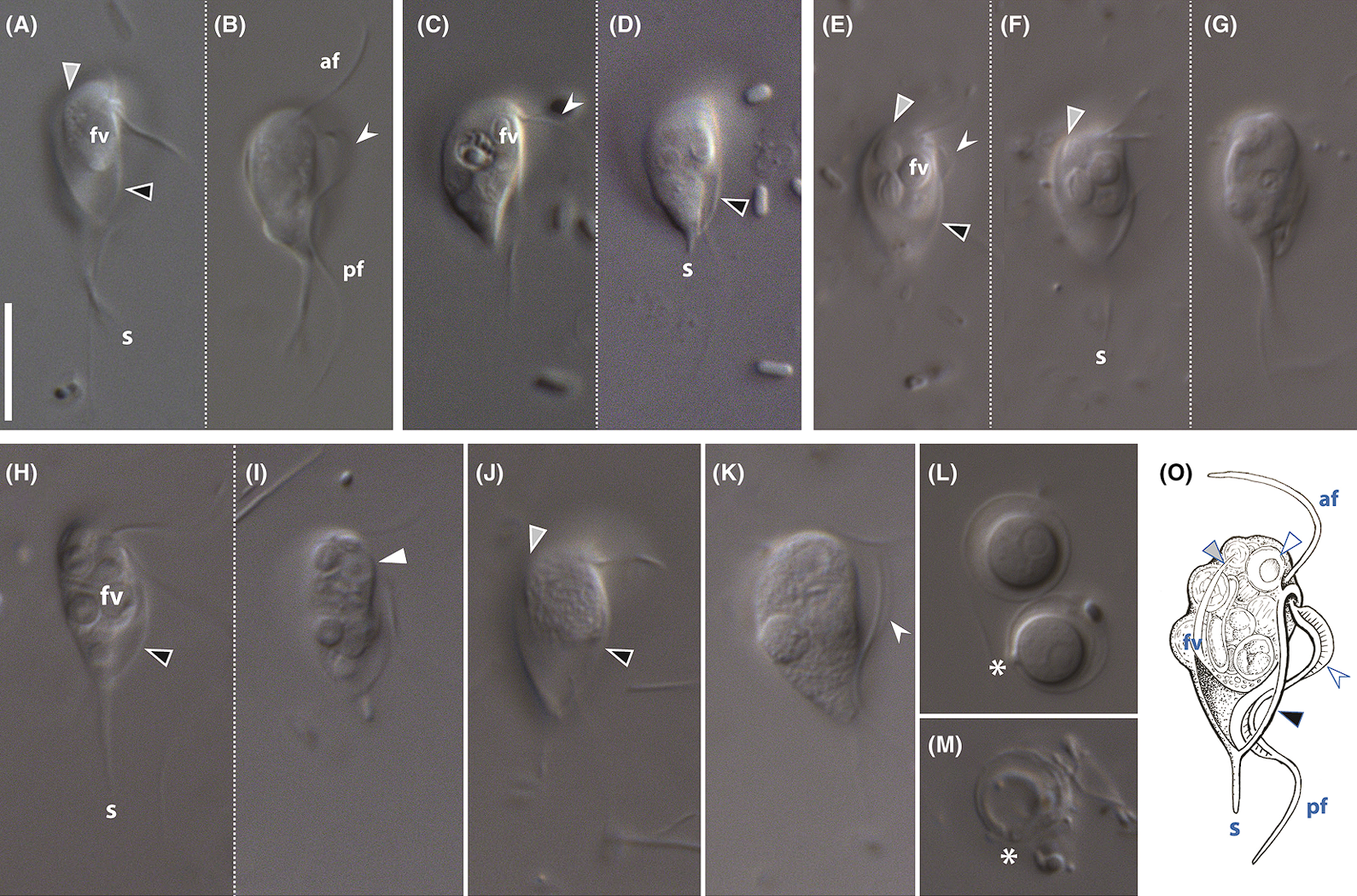
(上图:$S.~eglit$)
从形态上,驼背虫和巴塞尔虫除了鞭毛之外没啥相似之处,所以 $\text{BaSk}$ 的建立还是依靠分子生物学的证据。同时得出的另一项结论是,$\text{BaSk}$ 的姐妹群是后滴界第二大门——拱形虫门($\text{Fornicata}$)。
虽然因出产众多寄生虫而闻名,但拱形虫的系统发育树明白无误地告诉我们,它们的祖先也是自由生活的。如下图所示(系统发育未知的属没有标出):
位于最基部的颂天虫($\text{Carpediemonas}$)算是自由生活的拱形虫的代表。之所以叫这个名字,是因为它的一位发现者的妻子在那天去世了,于是就用格言“$Carpe~diem$”命名了这一物种,意为“采撷今日”。中文翻译可能是考虑到这句格言出自《颂歌集》,虽然意思差别有点大,但确实挺好听的。
言归正传,颂天虫生活在潮间带沉积物中,有前后两根鞭毛,前鞭毛负责运动,后鞭毛负责觅食。一些研究表明,颂天虫似乎可以“养殖”细菌,毕竟作为后滴虫,它无法进行完整的有氧代谢,排除的代谢废物刚好可以供细菌享用;等细菌吃的差不多了,再反过来吃掉一部分细菌,如此周而复始。
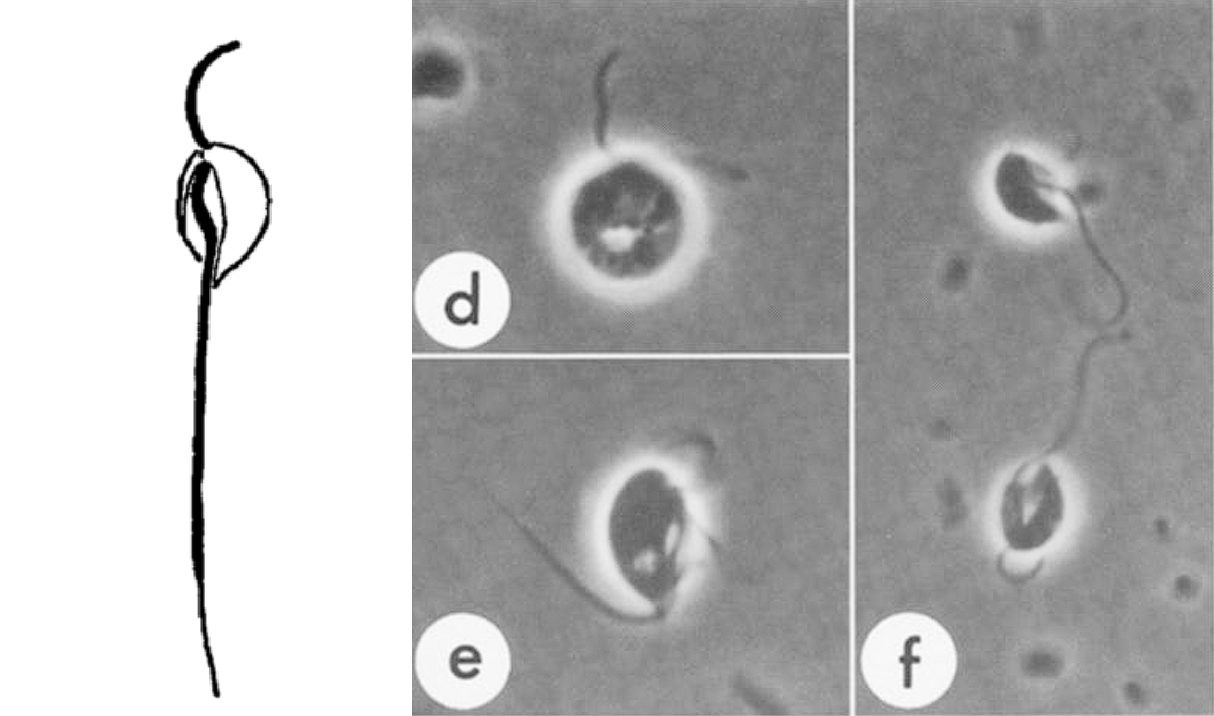
(上图:$C.~membranifera$ 的简笔画与显微图像)
一个令人困惑的事情是,颂天虫缺乏许多与 $\text{DNA}$ 复制相关的蛋白,也无法检测出编码这些蛋白质的基因。这种情况在后滴虫中是普遍的,但颂天虫非常极端。它是第一个如此剧烈地改动了普遍认为的 $\text{DNA}$ 复制和分离机制的真核生物,关于它是如何繁殖的,直到今天也只有几个语焉不详的假说;关于这种改动缘何而起,则更是难以回答。
在拱形虫门基部,有很多与颂天虫形态类似的物种,被统称为 $\text{CLOs}$。其中,中游虫($Hicanonectes$)似乎与颂天虫关系较近,此处被归入颂天虫科;其他的都被认为是独立的演化支。比如,$2016$ 年发现的腹钩虫($Aduncisulcus$)和 $2010$ 年发现的尽饮虫($Ergobibamus$)构成了腹钩虫纲($\text{Aduncisulcidea}$)。
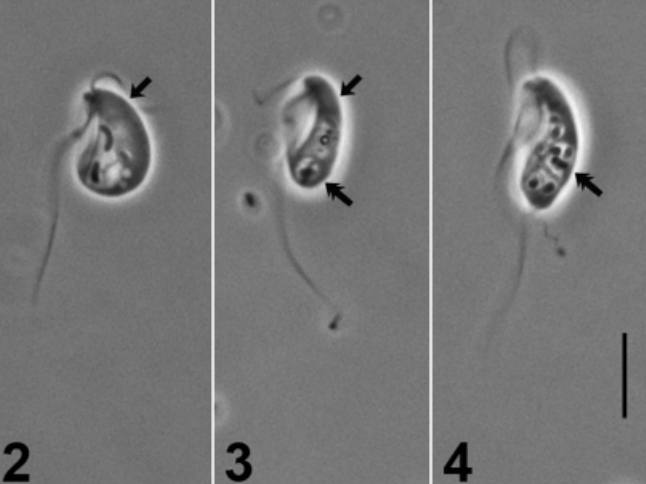
(上图:尽饮虫属的 $E.~cyprinoides$)
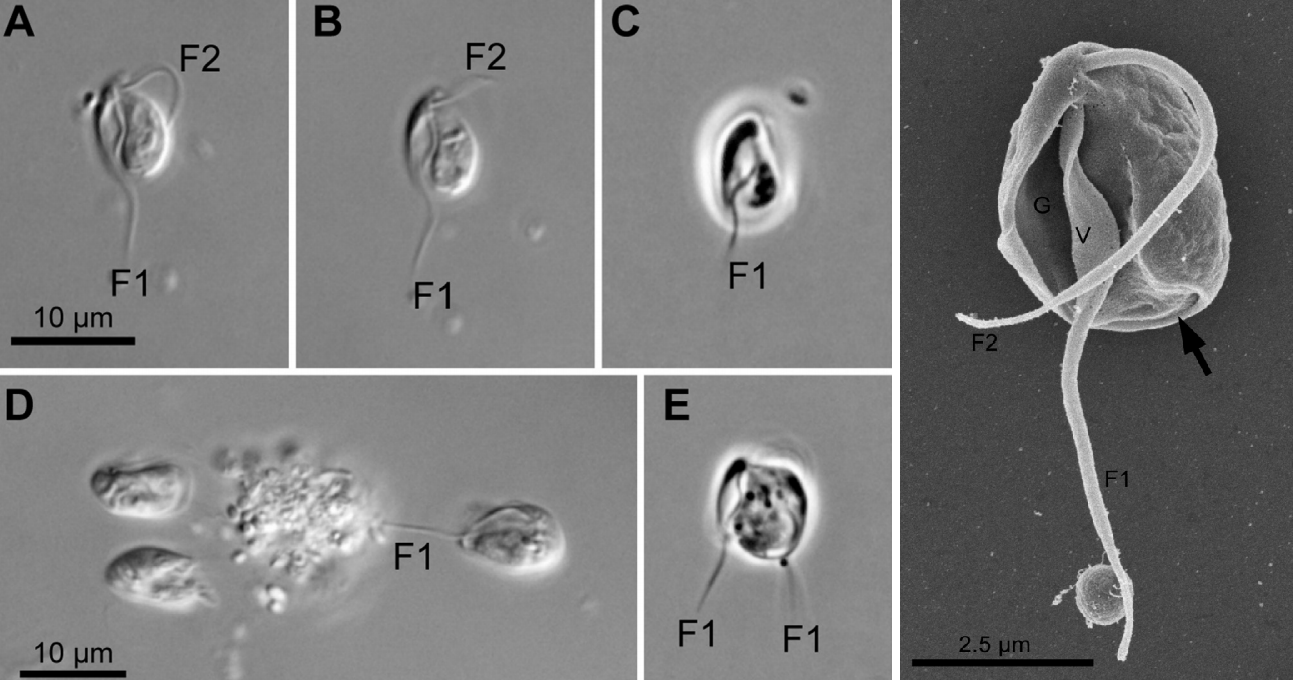
(上图:腹钩虫属的 $A.~paluster$)
唇鞭虫($Chilomastix$)曾被归入内滴虫目,但近几年的分子生物学研究将其独立为一纲。这个错误是难免的,因为属内四十多个物种中大部分是内生的,宿主以脊椎动物为多,且形态上确实有点向内滴虫那边靠的意思。最著名的莫过于寄生人类的迈氏唇鞭虫($C.~mesnili$)。它通过粪口途径传播,可能导致水样腹泻等症状;大部分物种一般被认为是肠道共生者而非寄生虫。
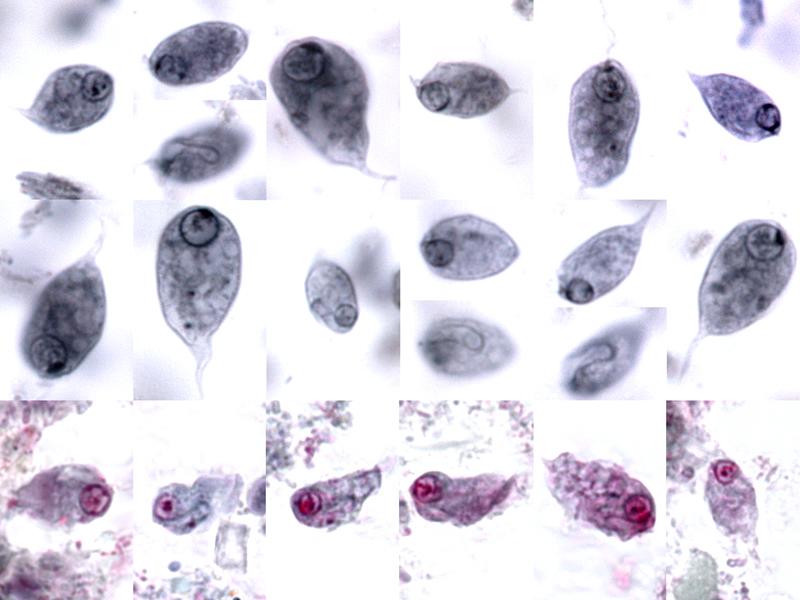
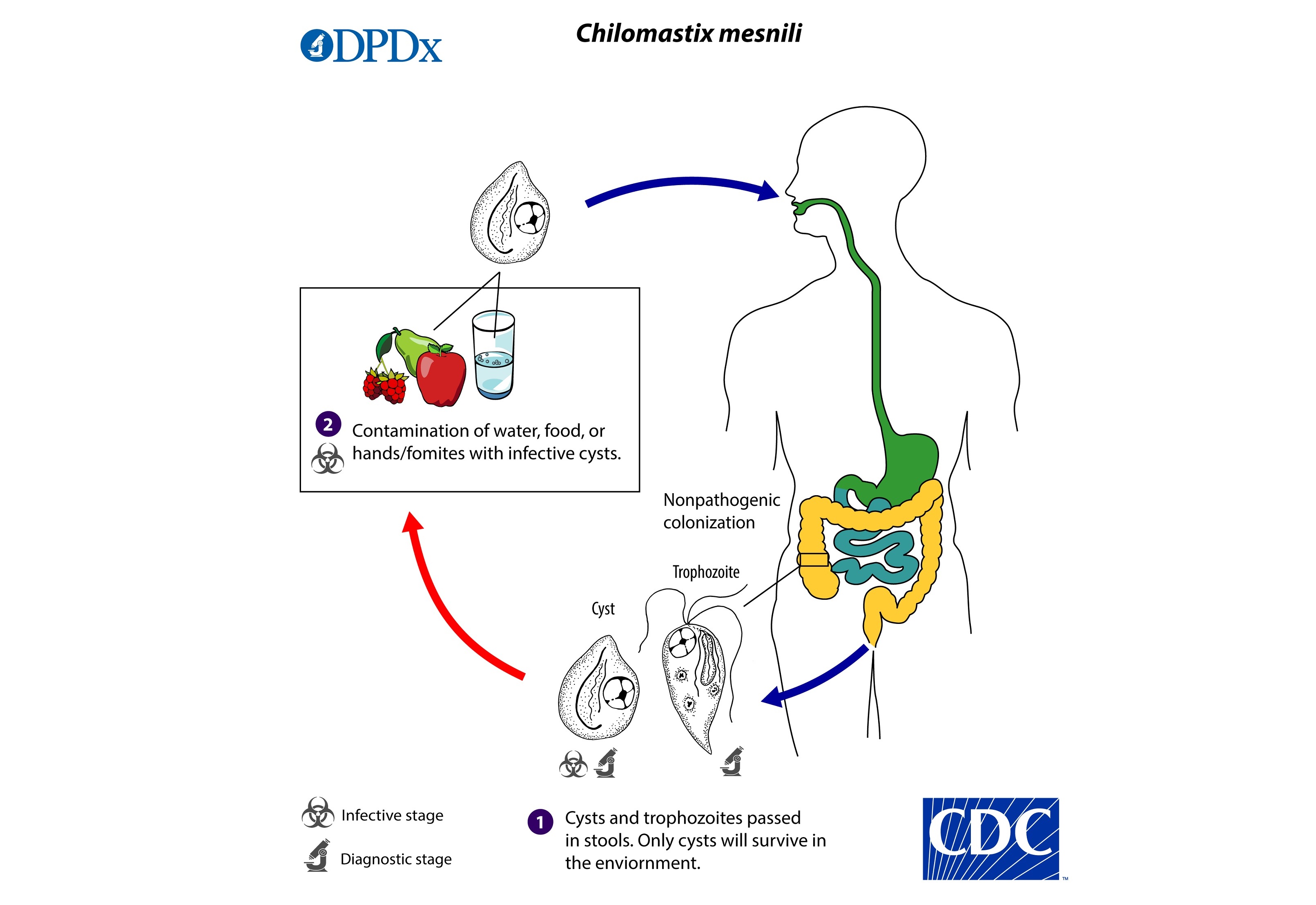
(上两图:$C.~mesnili$ 及其传播途径)
同属中也存在自由生活的物种,如 $C.~cuspidata$。与内生的种类相比,形态上并没有太大区别:
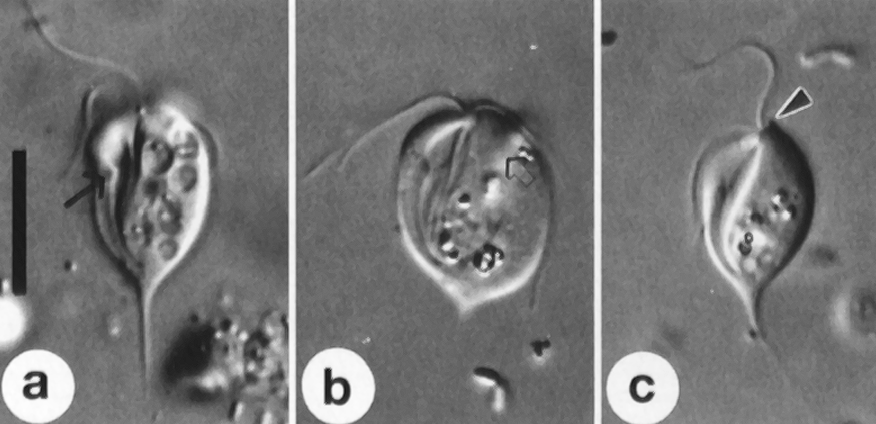
新月虫($Kipferlia$)在最新的研究中被认为是 $\text{CLOs}$ 中较深的类群,而不是处于基部。它有两根鞭毛,侧看像弯弯的新月,正看则像个葫芦。下图是趁它细胞分裂时拍的,所以看起来比较奇怪:
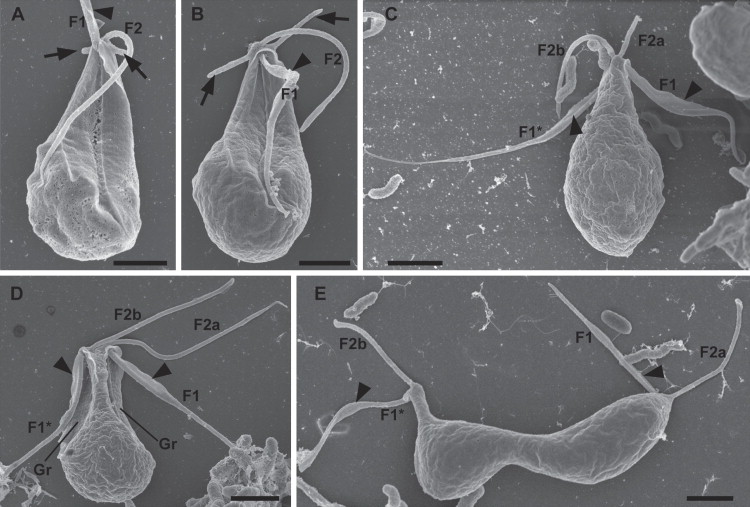
(上图:$K.~bialata$)
而新月虫的姐妹,则是拱形虫门中最大的一纲——胞咽虫纲($\text{Eopharyngia}$)。顾名思义,它们拥有一个叫胞咽($\text{cytopharynx}$)的结构,可以辅助它们进食(虽然在新月虫中也有类似的结构)。
胞咽虫纲的基群是双瓣虫($Dysnectes$),也是 $\text{CLOs}$ 中最深的类群,自由生活于海洋沉积物中:
(上图:$D.~brevis$)
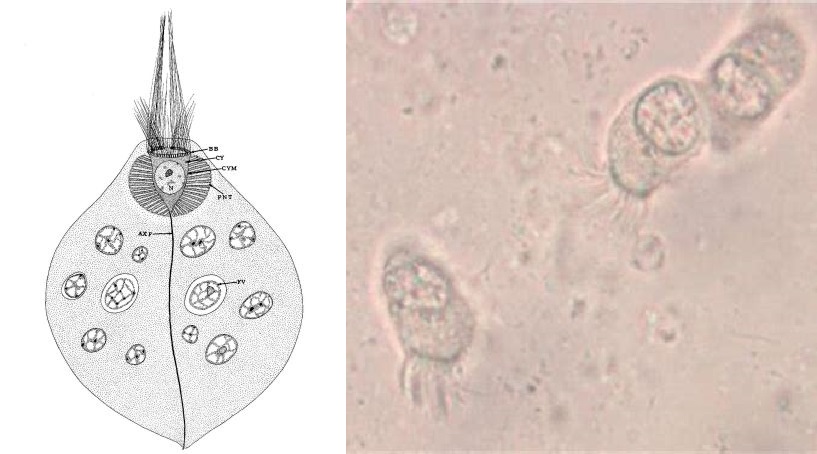
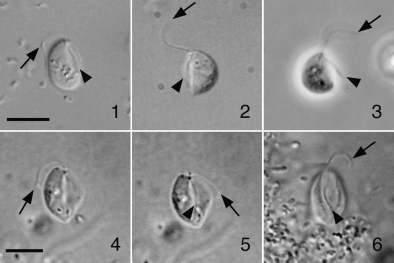
位于演化树更深处的内滴虫($Retortamonas$)虽然是内生的,但一般认为是无害的共生者。虽然也有一些临床病例,但没有强有力的证据支持内滴虫会导致疾病。最知名的物种是肠内滴虫($R.~intestinalis$)和中华内滴虫($R.~sinensis$),后者形成的包囊比前者大得多,因此可以区分;另一个区分点在它们的滋养体,后者的胞口大而明显,且鞭毛等长,而前者不等长(虽然在简笔图上看不出来)。
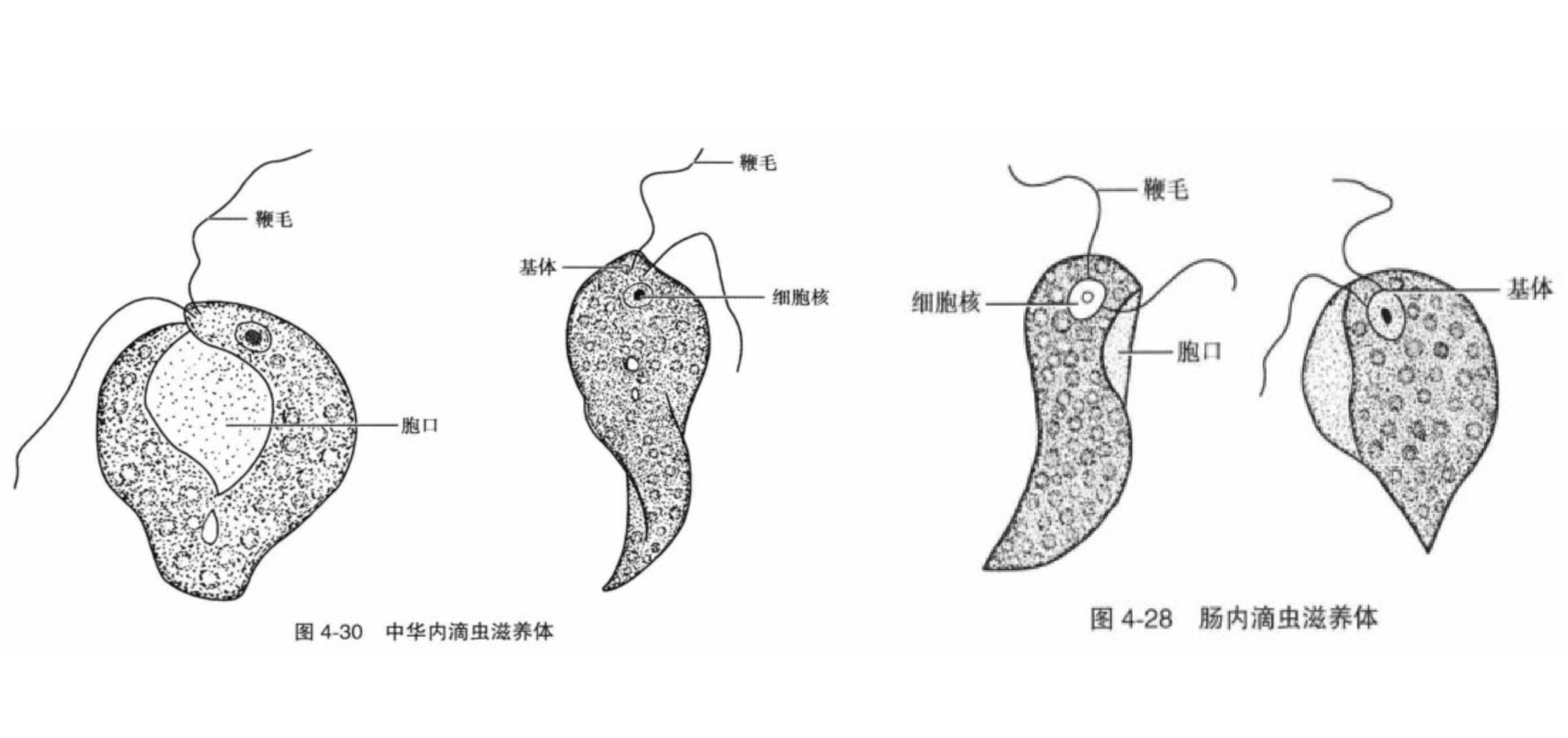
(上图:$R.~intestinalis$ 和 $R.~sinensis$ 的简笔画)
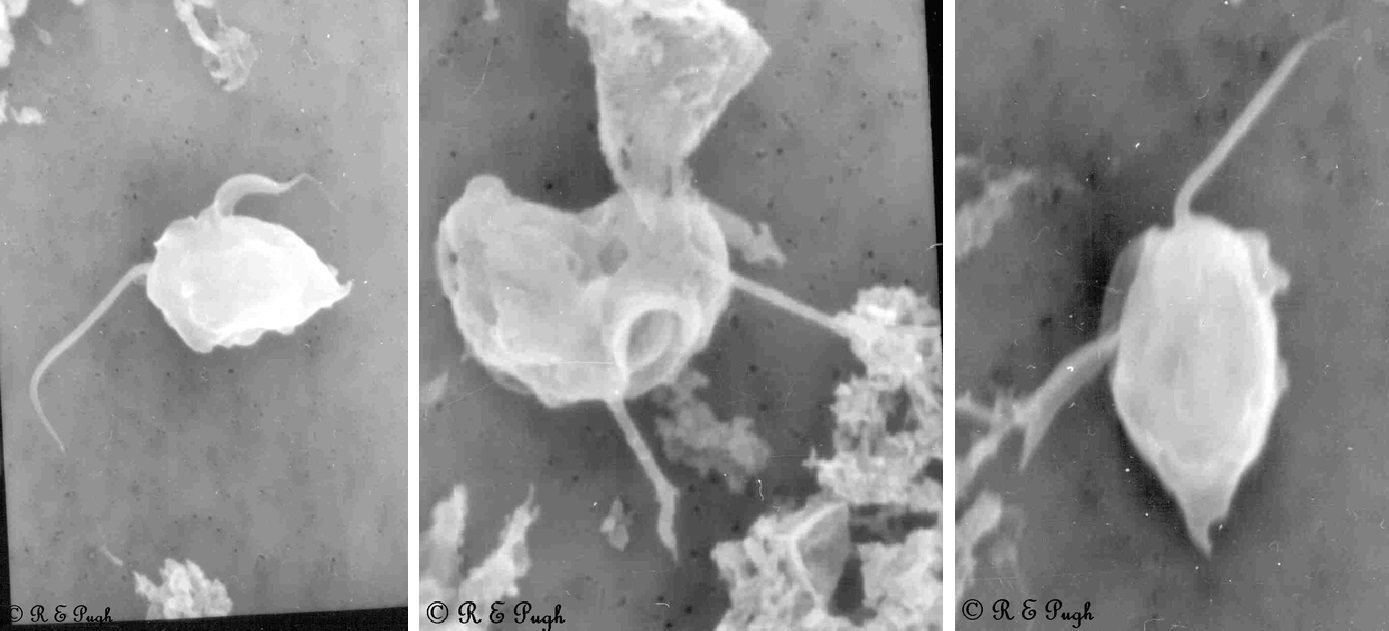
(上图:$R.~intestinalis$ 实拍)
相比之下,双滴目($\text{Diplomonadida}$)下的成员就没那么老实了。虫如其名,双滴目形态上最标志性的特征,正是其鞭毛虫形态有两个核,且基本两侧对称。虽然不太清楚这样的身体模式究竟有什么用,但毫无疑问的是,双滴目是整个拱形虫门的排面所在,很多大名鼎鼎的寄生虫都来自这个类群。
贾第鞭毛虫属($Giardia$)是其下最大的一属,已知物种超过三十个,全部寄生于脊椎动物体内。唯一一种寄生人类的,就是著名的蓝氏贾第鞭毛虫($G.~lamblia$,有许多异名)。它无疑是影响人类最广泛的肠道寄生虫之一,在卫生条件较差的国家更甚,一些地区的感染率甚至可以高达 $33\%$。与内滴虫不同,$G.~lamblia$ 会显著扰乱正常的营养吸收,它的滋养体会分泌有毒物质诱导细胞凋亡,还能通过一系列手段规避免疫反应。
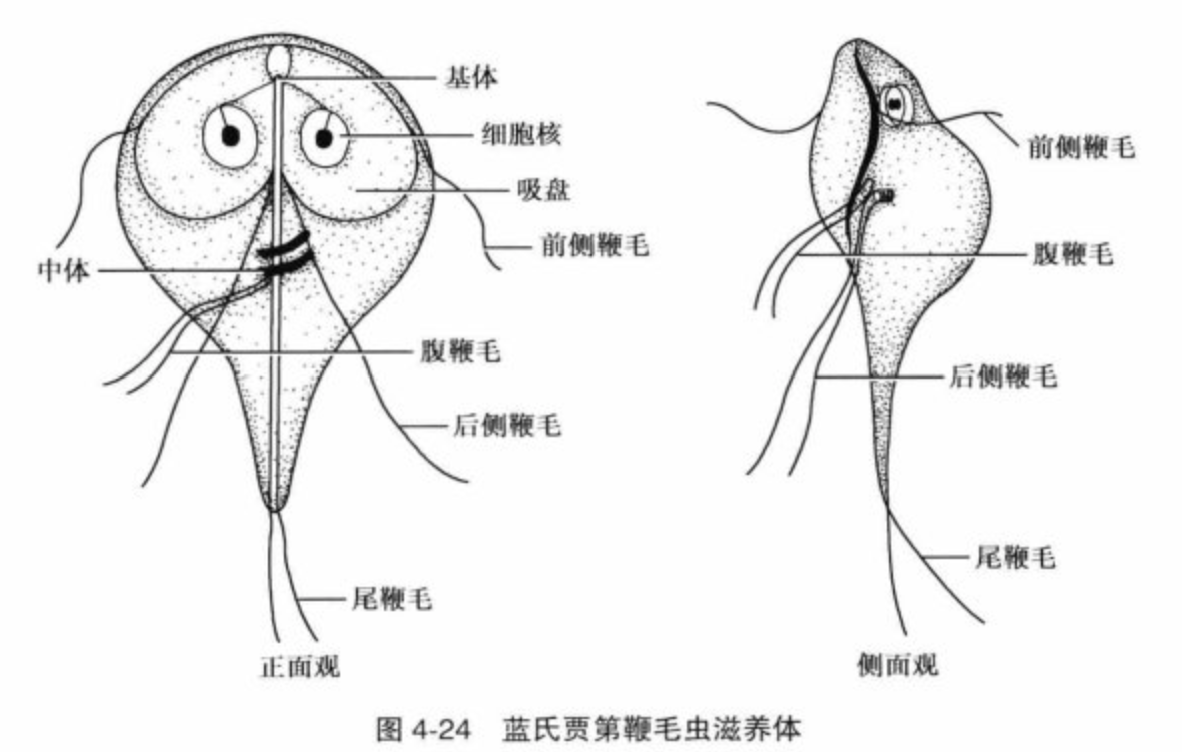
(上图:$G.~lamblia$ 的滋养体结构)
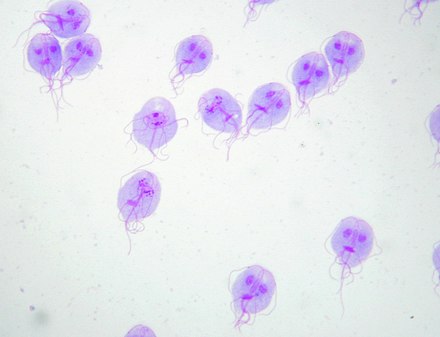
(上图:$G.~lamblia$ 的染色实拍)
它的所作所为会导致严重的腹痛、腹泻,并可能导致体内电解质大量流失,进而导致神经信号紊乱等严重后果。虽然可以医治,但由于一些国家在此方面的落后,$G.~lamblia$ 每年依然会导致上万人死亡。对于儿童来说,感染 $G.~lamblia$ 会导致发育迟缓,从而造成终身性的影响。不过对于 $G.~lamblia$ 来说这非常不错,它将巨量的包囊通过感染者的粪便释放到环境中(每个感染者每天会排出百万到十亿个包囊),这些包囊可以休眠数月,直到被新的宿主摄入。因此,它传染性极强,加之可以人畜共患,使得它在世界范围内成为了重大的公共卫生问题。
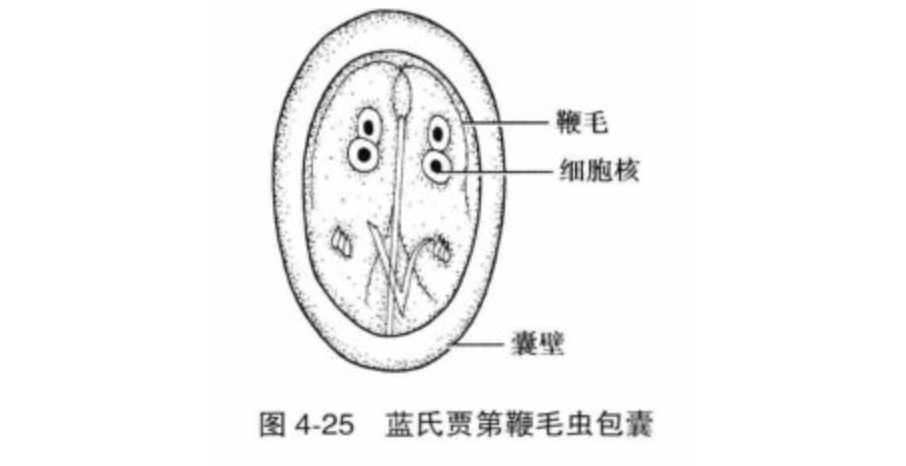
(上图:$G.~lamblia$ 的包囊简笔。成熟的包囊有四个细胞核,且囊壁很厚)
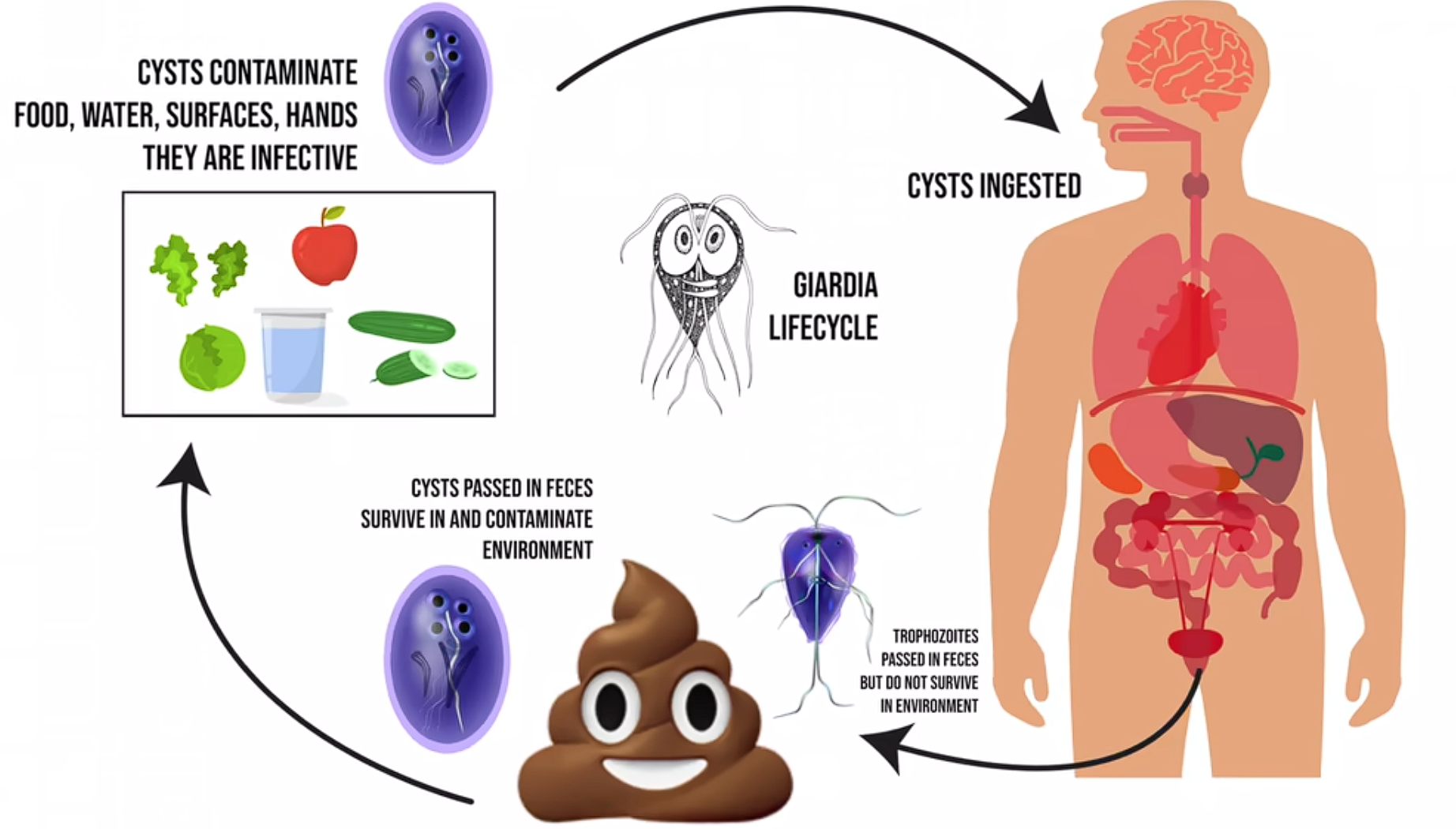
(上图:$G.~lamblia$ 的生命历程示意)
一个通俗化讲解 $G.~lamblia$ 感染病例的视频:Link
与贾第鞭毛虫属同属贾第虫科($\text{Giardiinae}$)的八鞭毛虫属($Octomitus$)则主要感染啮齿目哺乳动物,二者形态上很相似:
(上图:$O.~intestinalis$)
双滴虫的另一支,六前鞭毛虫科($\text{Hexamitidae}$)则是一个非常大的类群。它们的形态共同点就写在名字里,多数种类有三对(六条)向前的鞭毛。比较有意思的是,该科中存在自由生活的类群。转滴虫属($Trepomonas$)是其中的代表,事实上,它也是科中最早被发现的类群。
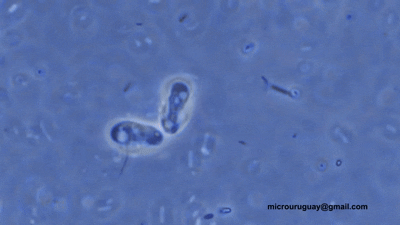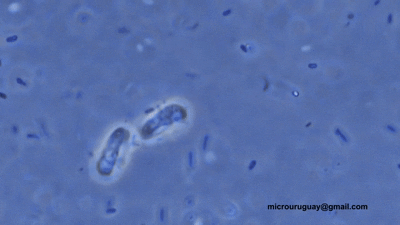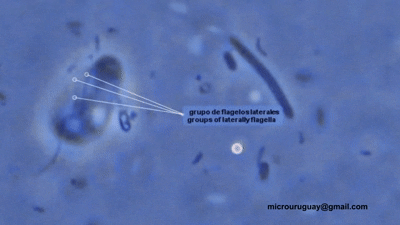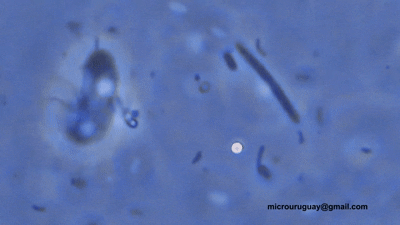
(上动图:$T.~agilis$)
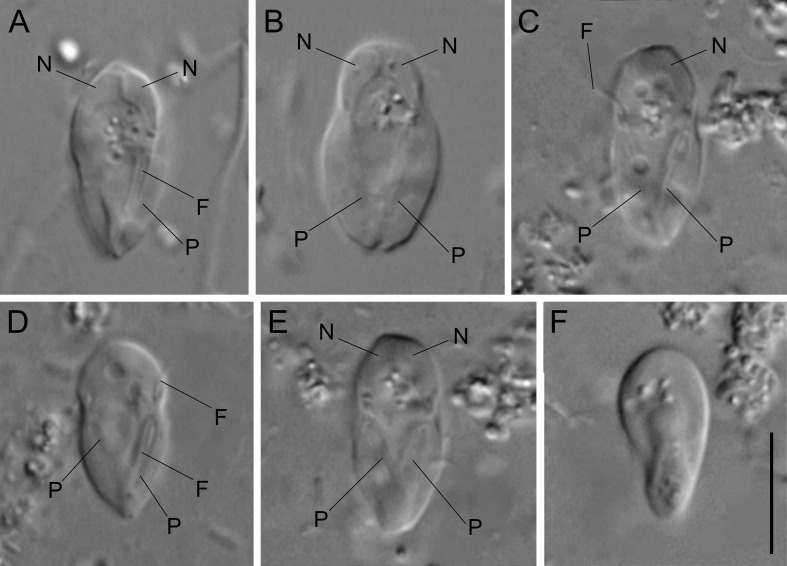
(上图:$T.~latecapitata$)
旋胞虫属($Gyromonas$)是双滴目中体型最小的类群,因此比较难拍,但可以确定它也是自由生活的。它似乎有一个小的伪足,形状也比同科的其他类群更尖一些,细胞核隐约可见:
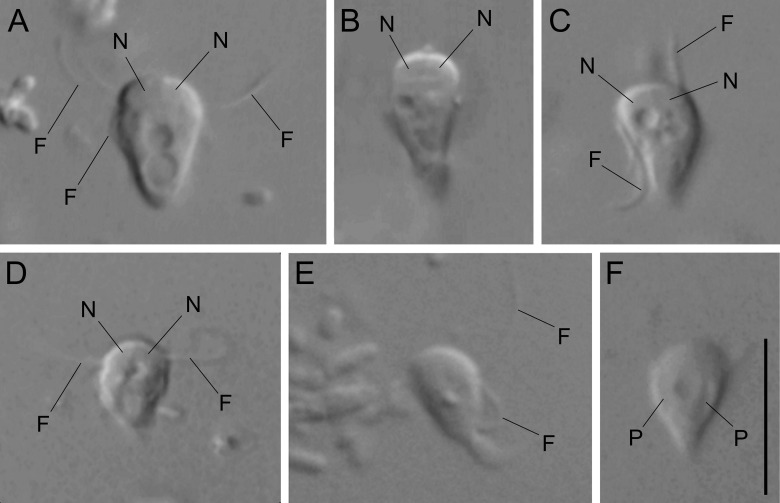
(上图:$G.~ambulans$)
话说回来,六前鞭毛虫中寄生的类群还是占了大头。但且慢,让我们再来看看它的系统发育。按照常理,生物向寄生生活的转变是不可逆的,因此一个类群如果同时有寄生和自由生活的子类群的话,自由类群的位置一般在演化树基部。先前我们的讨论的类群也基本都遵守这一点,自然地,人们认为转滴虫和旋胞虫也应当位于基部。但是,几乎所有分子生物学证据都得出了截然相反的结论。
让我们再画一遍胞咽虫纲的系统发育树,但将内生的类群用红色标记,自由的类群用绿色标记:
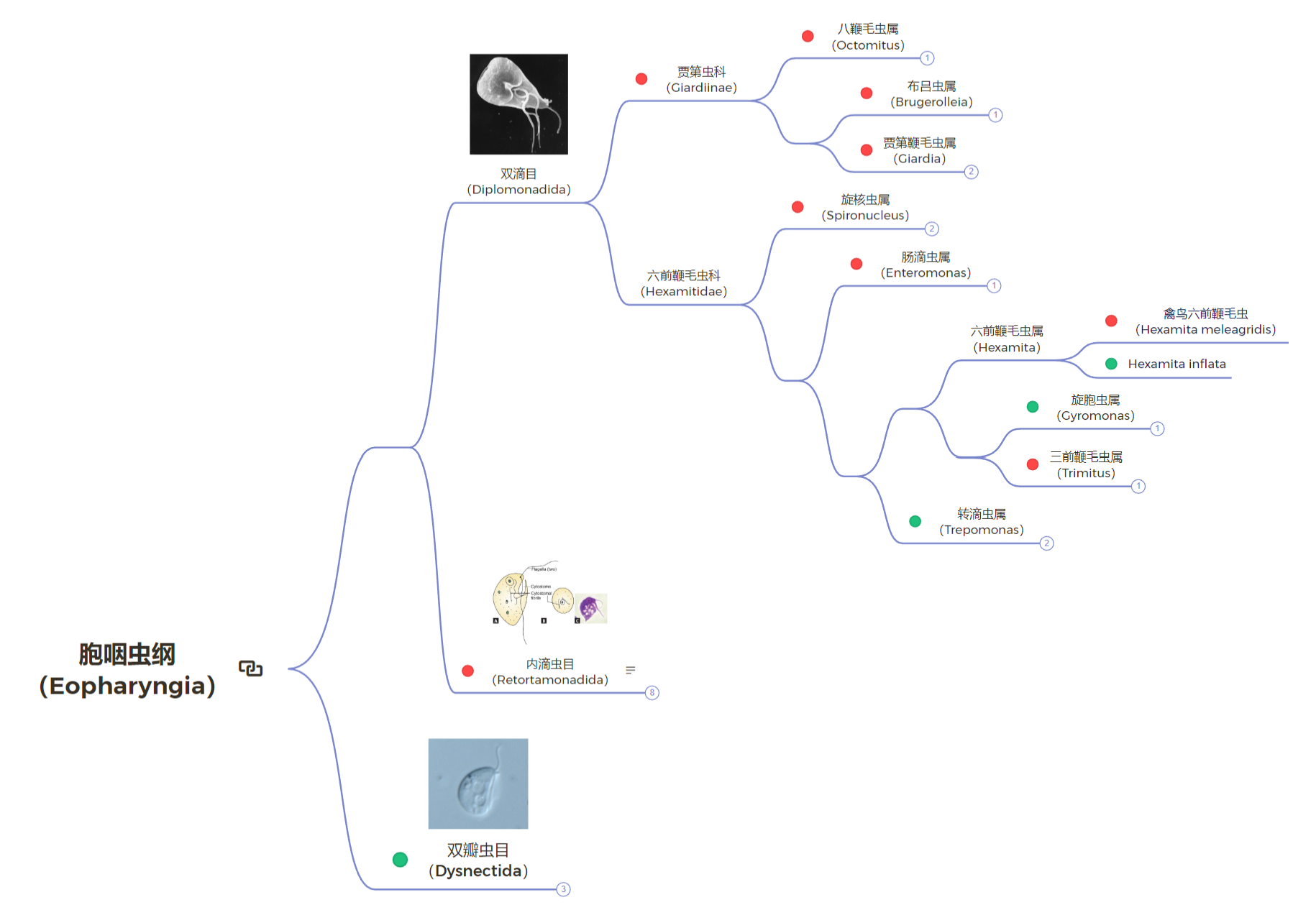
需要注意的是,六前鞭毛虫科中有很多类群系统发育位置是位置的,因此没有在上图中呈现。可以看到,自由类群反而出现在了演化树的深处。更进一步,六前鞭毛虫属($Hexamita$)中既有自由生活的物种 $H.~inflata$,也有著名的寄生物种禽鸟六前鞭毛虫($H.~meleagridis$)。虽然该属中各物种之间的关系很不明确(上图中物种也不全),但已经可以说明很多问题。
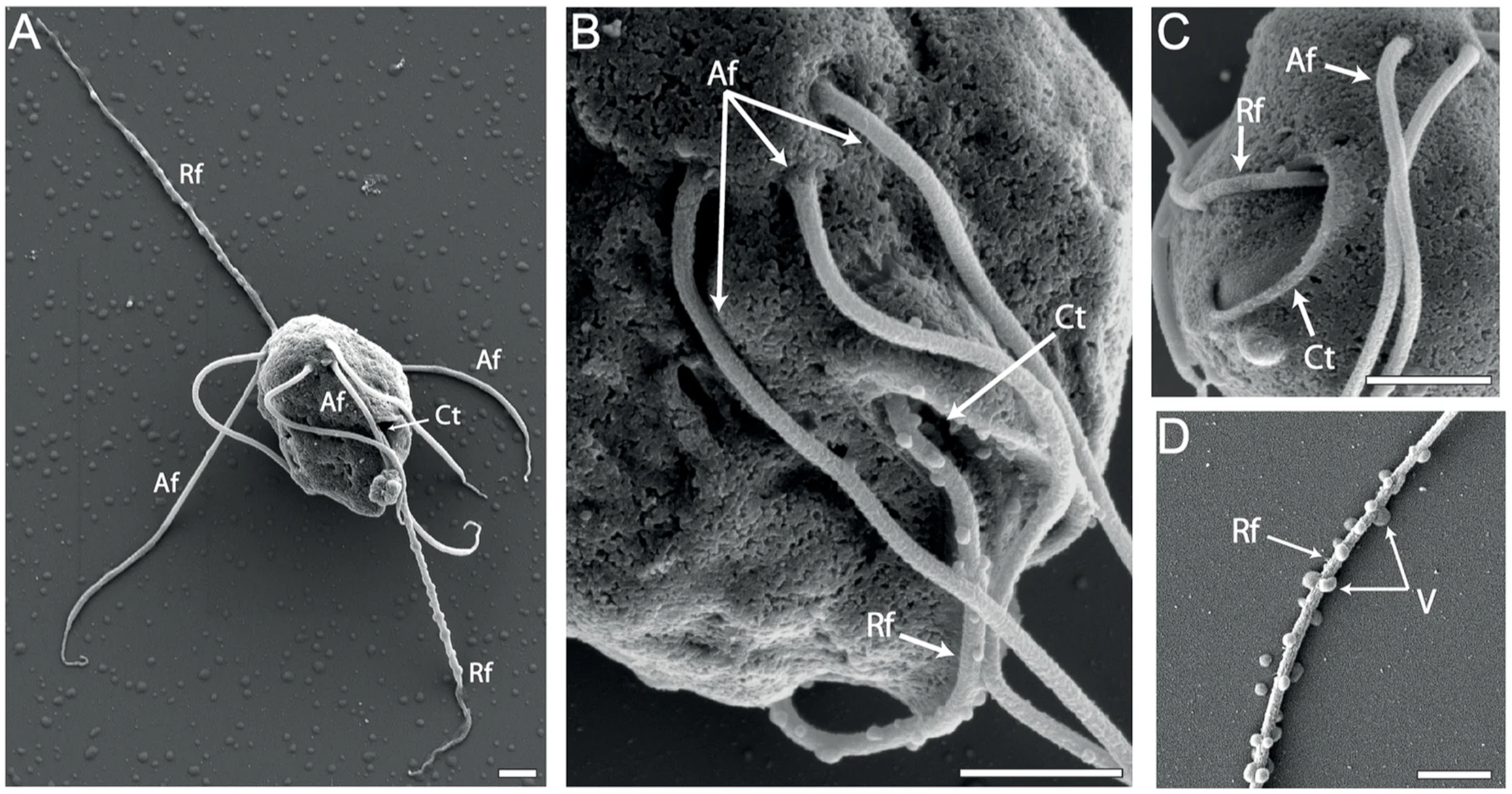
(上图:$H.~inflata$ 的超微结构摄影)
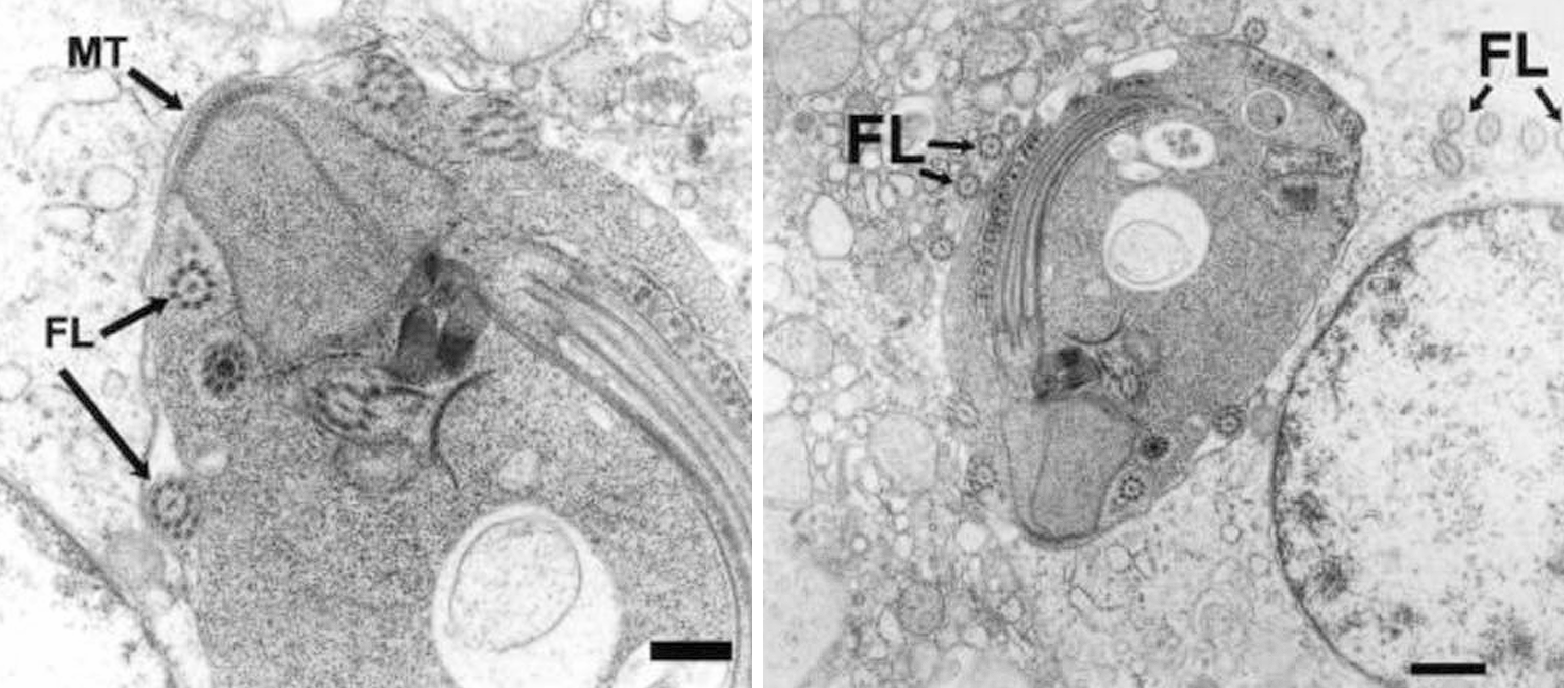
(上图:寄生于鸟类组织中的 $H.~meleagridis$)
如果我们仍然坚持之前的理论,那么不可避免的,仅双滴目中就至少有五次独立的由自由向内生的转变,算上系统发育位置不确定的只会更多。结合一些其他证据,有人提出了这样一个假说:整个双滴目起源于同一个内生祖先,而那些自由生活的类群,祖上也是内生的。但这又带来了另一个问题——它们是如何做到的呢?
一个演化支向内生转变,往往意味着它们会失去一些对自由生活而言非常重要的代谢途径,同时会在形态上发生较大变化以适应。但对于双滴虫而言,至少后面这条不太成立。作为单细胞生物(虽然有两个核),它们的身体模式简单但通用,因此并不需要太多的转变。就行上面看到的 $H.~inflata$ 和 $H.~meleagridis$ 基本只能靠周围环境区分,因为它们的身体结构相差无几(不然怎么被分到一个属里)。至于代谢问题,我们目前则仍未完全搞清楚。
我们同时也注意到,自由生活的双滴虫也不是单系的,说明回归自由的转变发生了不止一次。但是,它们连同一些内生的亲戚在一起,可以组成六前鞭毛虫科中的一个单系群(六前鞭毛虫亚科,未被正式承认)。或许是该类群的共同祖先未能完全特化,比如它可以在自由和内生之间灵活转变,甚至同时有自由和内生两个生命周期,即双生($\text{amphizoic}$)。若真是这样,那它的部分后代抛弃掉内生生活就变得合理了。这也并非毫无根据,因为在其他古虫类群如渗养门($\text{Percolozoa}$)中,我们确实发现了这样的物种,比如著名的内格里虫属($Neagleria$):
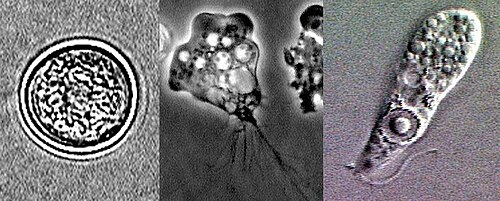
(上图:不同形态的 $N.~fowleri$,属渗养门,是双生物种的代表)
事实上,由于我们对双滴虫仍然缺乏了解,很多物种的生命周期都存在空白,其中就包含一些潜在的双生物种。如果真的证实了双生物种的存在,那无疑将成为该假说的一个重要支撑。至于究竟有没有,就让未来的研究告诉我们答案吧。
3. 后滴虫演化位置的争议
文章的末尾,让我们来讨论一下在开头留下的问题。实话实说,后滴虫真的如同幽灵一般,难以确定其演化位置。截至我写文章的这一刻,似乎还有三种主流的观点,让我们一一道来。
第一种是传统观点,认为后滴虫分化在真核生物两大类群分开之后,且更接近槽虫类,也就是包括动物、真菌在内的演化支(以此可以定义泛槽虫类)。当然,它认为后滴界是单系的:
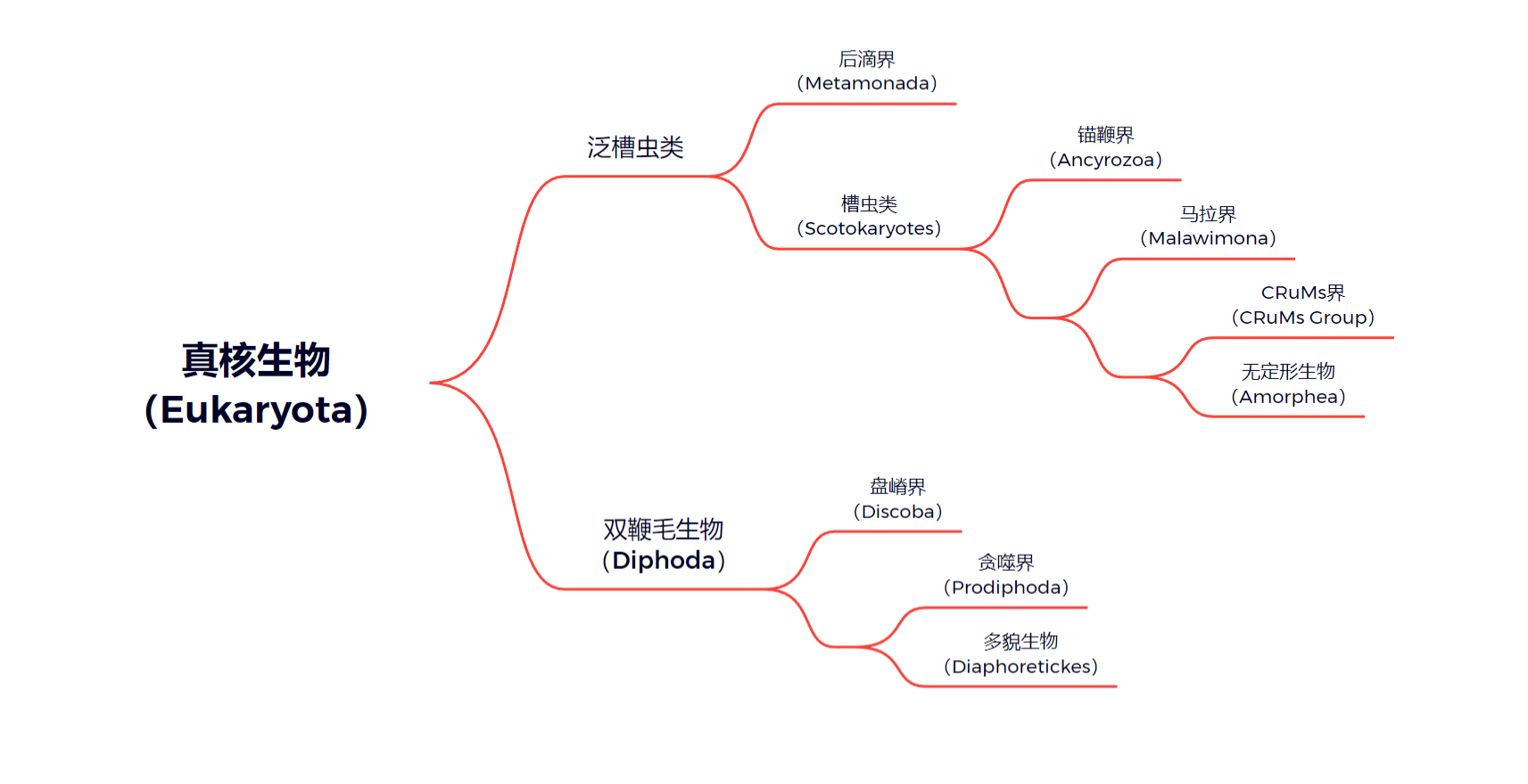
第二种可以称为半激进观点,认为后滴界应是真核生物最基部的分类群。此观点定义了新核生物这一演化支,是后滴界的姐妹,包含槽虫类和双鞭毛生物。后滴界本身保持单系性:
第三种更为激进。在第二种观点的基础上,它不再承认后滴虫的单系性,认为它高度并系;持有这一观点的人通常也认为盘嵴界($\text{Discoba}$)应处于更基部的位置,而不是双鞭毛生物中。此观点进一步附加了正核生物这一演化支:
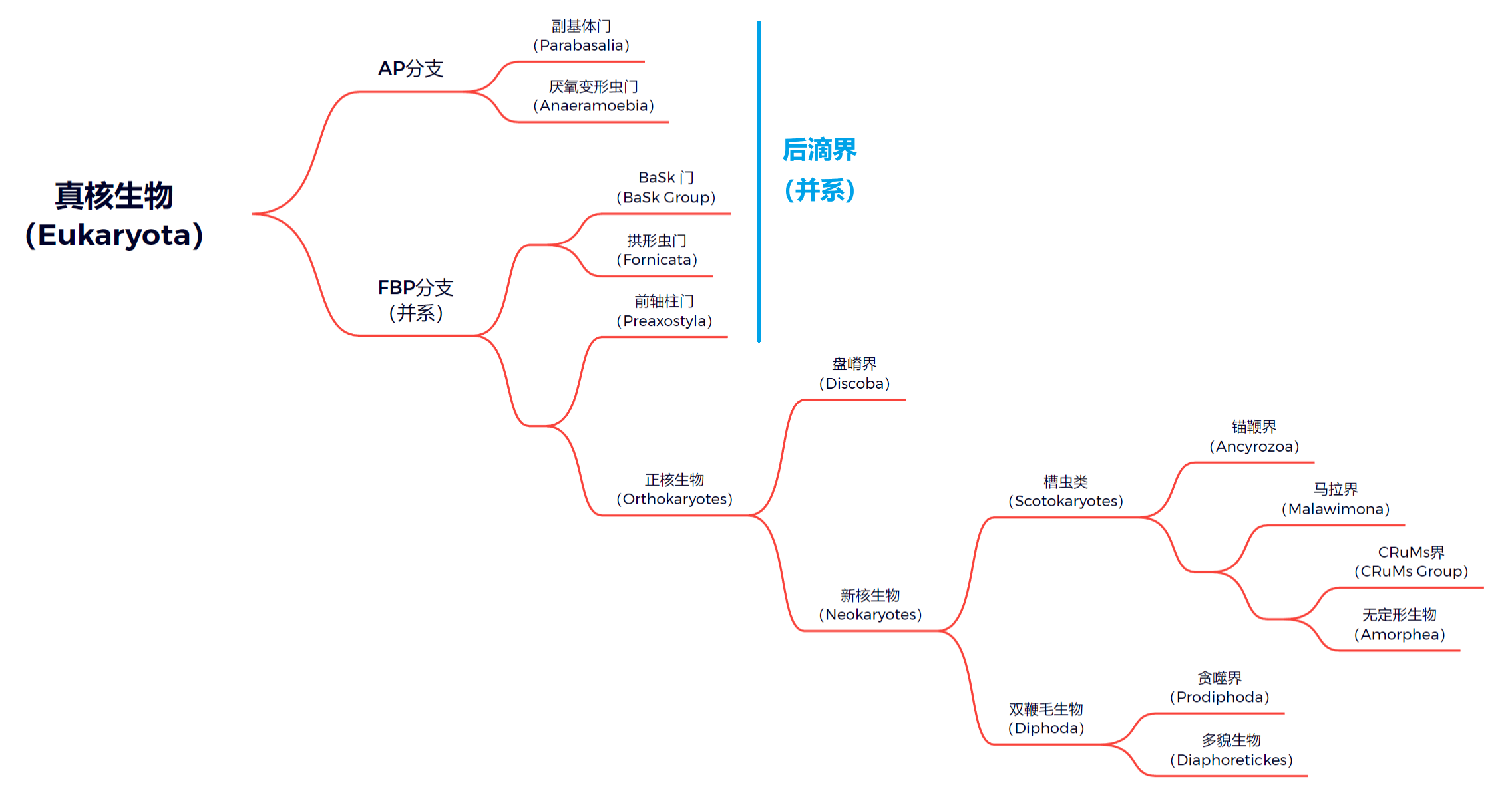
当然,对于槽虫类和双鞭毛生物内部类群的演化位置也有争议,但这不是我们讨论的重点。
我们可以看到,第三种观点与传统演化理论是严重相悖的。传统认为,后滴虫并非从来就没有线粒体,而是它们线粒体的有氧呼吸功能退化了,并被进一步改造成了后滴虫特色的氢小体等细胞器。至于为何其中会检测出 $δ/γ-$变形菌的基因,那就是在共同祖先那儿发生了一次水平基因转移罢了,这种事件在原生生物那儿屡见不鲜,线粒体退化也可以找到类似的例子;在后滴界内部,也可以找到一些证据,比如前文已经讨论过的关于单轴柱虫的争论。
但是,如果后滴虫是并系的话,则至少有三个演化支都“独立”地退化了线粒体的有氧呼吸,“独立”地把它改造为氢小体,“独立”地发生了同一类细菌的水平基因转移······这显然很不合理。因此,该观点的支持者提出了新的演化假说,即连续内共生假说。
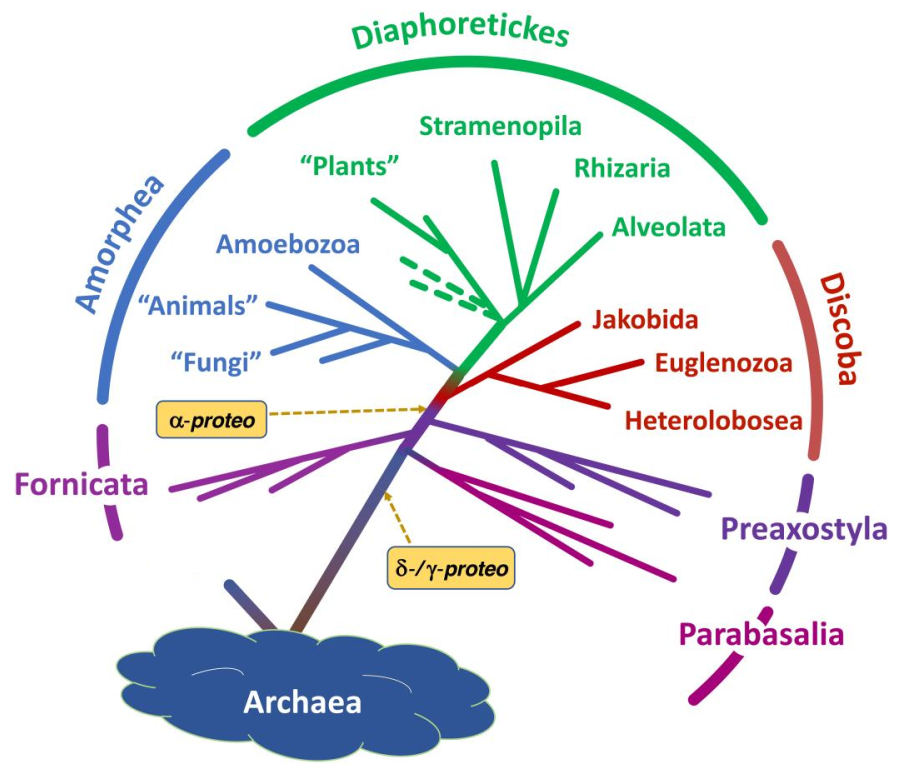
简而言之,该假说认为在真核生物演化早期发生了两次内共生事件:第一次内共生是 $δ/γ-$变形菌的内共生,这造就了后滴虫的氢小体等细胞器;在第二次内共生之前,后滴虫三大类群先后分化,最晚分化的是前轴柱门;第二次内共生才轮到 $α-$变形菌,此次内共生造就了真正的线粒体。不论是直接取代了氢小体还是利用自己的有氧呼吸能力改造了它,总之线粒体才是后来者。这一假说可谓石破天惊,因为它直接改写了真核生物早期演化的图景。
当然,该假说也并非没有证据。首先,后滴界之外的真核生物虽说都有真正的线粒体,但线粒体中还是可以检测到 $γ-$变形菌的基因,尽管比 $α-$变形菌少;其次,另一类古虫——盘嵴界之所以叫这个名字,是因为它们拥有独一无二的盘状线粒体嵴,而在这一假说中,真核生物有氧呼吸的起源较晚,一直要拖到正核生物的最近共祖那里,而盘嵴界被认为是正核生物的基干分支,这有助于解释这一现象。此外还有很多基因层面的证据,但事实上,这些证据都不是强有力的、可以一锤定音的那种,反驳者也可以找到自己的说辞。
那么,到底哪一种说法是正确的呢?或者说,没有一种是正确的,后滴虫的演化远比我们所提出的一切假说都更加复杂。相信你已经有了自己的答案。希望有朝一日,我们终可以破解这一谜团,后滴虫也不再是演化树上的那个永远的幽灵。
4. 结语
这是我第一次写生物方面的科普长文,此前没什么经验,只是一个爱好者的心血来潮罢了,还请各位读者多多包涵。如果你发现文章有任何错误,敬请指出,在此感谢!(当然,这并不意味着我要转行,化学方面的内容日后也会更新的,敬请期待)
特别致谢:@博养万物(贴吧同名)大佬,它为这篇文章,尤其是系统发育树提供了许多建议;在与他的交流中,我也学到了许多。
最后,贴上一张完整的,后滴虫从界到属的系统发育树:
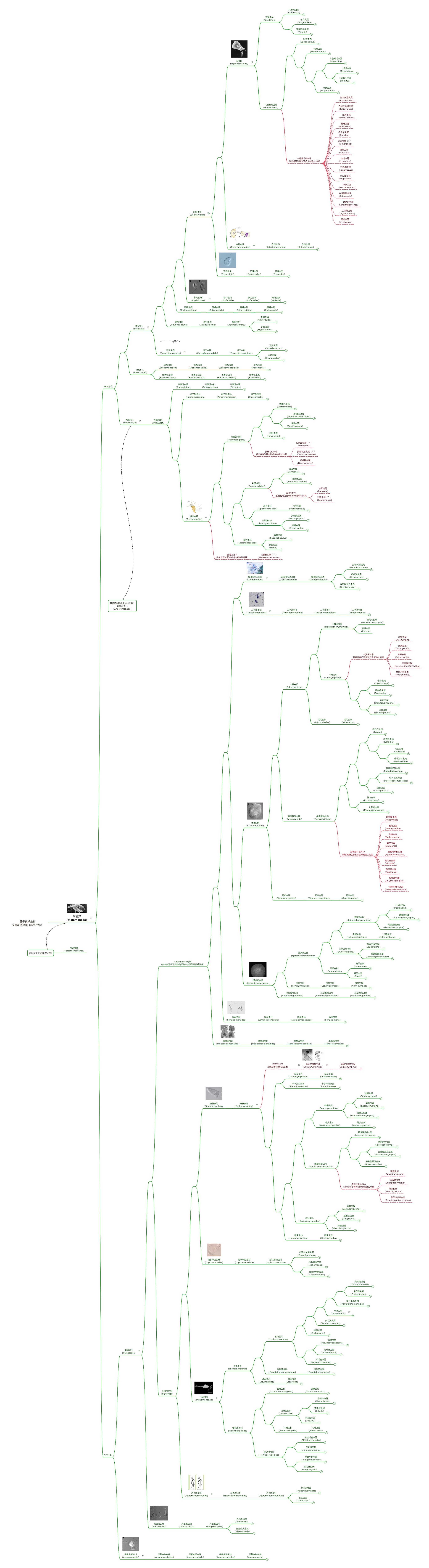
以及它的 $\text{pdf}$ 版下载链接:Link
5.参考文献
$[1]$:Williams, S.K., Jerlström Hultqvist, J., Eglit, Y. et al. Extreme mitochondrial reduction in a novel group of free-living metamonads. Nat Commun 15, 6805 (2024).
$[2]$:Táborský P, Pánek T, Čepička I. Anaeramoebidae fam. nov., a novel lineage of anaerobic amoebae and amoeboflagellates of uncertain phylogenetic position[J]. Protist, 2017, 168(5): 495-526.
$[3]$:Boscaro V, James E R, Fiorito R, et al. Updated classification of the phylum Parabasalia[J]. Journal of Eukaryotic Microbiology, 2024, 71(4): e13035.
$[4]$:Céza V, Kotyk M, Kubánková A, et al. Free-living trichomonads are unexpectedly diverse[J]. Protist, 2022, 173(4): 125883.
$[5]$:Céza V, Pánek T, Smejkalová P, et al. Molecular and morphological diversity of the genus Hypotrichomonas (Parabasalia: Hypotrichomonadida), with descriptions of six new species[J]. European journal of protistology, 2015, 51(2): 158-172.
$[6]$:Cepicka I, Dolan M F, Gile G. Parabasalia[M]//Handbook of the Protists: Second Edition. Springer International Publishing, 2017: 1175-1218.
$[7]$:Hehenberger E, Boscaro V, James E R, et al. New Parabasalia symbionts Snyderella spp. and Daimonympha gen. nov. from South American Rugitermes termites and the parallel evolution of a cell with a rotating “head”[J]. Journal of Eukaryotic Microbiology, 2023, 70(5): e12987.
$[8]$:Poinar Jr G. Early cretaceous protist flagellates (Parabasalia: Hypermastigia: Oxymonada) of cockroaches (Insecta: Blattaria) in Burmese amber[J]. Cretaceous Research, 2009, 30(5): 1066-1072.
$[9]$:Al Jewari C, Baldauf S L. An excavate root for the eukaryote tree of life[J]. Science Advances, 2023, 9(17): eade4973.
$[10]$:Novák L V F, Treitli S C, Pyrih J, et al. Genomics of preaxostyla flagellates illuminates the path towards the loss of mitochondria[J]. PLoS Genetics, 2023, 19(12): e1011050.
$[11]$:Karnkowska A, Vacek V, Zubáčová Z, et al. A eukaryote without a mitochondrial organelle[J]. Current Biology, 2016, 26(10): 1274-1284.
$[12]$: Yazaki E et al. 2020 Barthelonids represent a deep-branching metamonad clade with mitochondrion-related organelles predicted to generate no ATP. Proc. R. Soc. B 287: 20201538.
$[13]$:Catherine Bernard, Alastair G. B. Simpson & David J. Patterson (2000) Some free-living flagellates (protista) from anoxic habitats, Ophelia, 52:2, 113-142, DOI: 10.1080/00785236.1999.10409422
$[14]$:Eglit Y, Williams S K, Roger A J, et al. Characterization of Skoliomonas gen. nov., a haloalkaliphilic anaerobe related to barthelonids (Metamonada)[J]. Journal of Eukaryotic Microbiology, 2024, 71(6): e13048.
$[15]$:Ekebom J, Patterson D J, Vørs N. Heterotrophic flagellates from coral reef sediments (Great Barrier Reef, Australia)[J]. Archiv für Protistenkunde, 1996, 146(3-4): 251-272.
$[16]$:Park J S, Kolisko M, Simpson A G B. Cell morphology and formal description of Ergobibamus cyprinoides ng, n. sp., another Carpediemonas‐like relative of diplomonads[J]. Journal of Eukaryotic Microbiology, 2010, 57(6): 520-528.
$[17]$:Yubuki N, Huang S S C, Leander B S. Comparative Ultrastructure of Fornicate Excavates, Including a Novel Free-living Relative of Diplomonads: Aduncisulcus paluster gen. et sp. nov[J]. Protist, 2016, 167(6): 584-596.
$[18]$:Wiśniewska M M, Salomaki E D, Silberman J D, et al. Expanded gene and taxon sampling of diplomonads shows multiple switches to parasitic and free-living lifestyle[J]. BMC biology, 2024, 22(1): 217.
$[19]$:Yubuki N, Inagaki Y, Nakayama T, et al. Ultrastructure and ribosomal RNA phylogeny of the free‐living heterotrophic flagellate Dysnectes brevis n. gen., n. sp., a new member of the Fornicata[J]. Journal of Eukaryotic Microbiology, 2007, 54(2): 191-200.
$[20]$:Mazancová E, Zadrobílková E, Yubuki N, et al. Phylogenetic and morphological diversity of free-living diplomonads[J]. European Journal of Protistology, 2023, 91: 126024.
$[21]$:Akdeniz Z, Havelka M, Stoklasa M, et al. The expanded genome of Hexamita inflata, a free-living diplomonad[J]. Scientific Data, 2025, 12(1): 192.
$[22]$:Cooper G L, Charlton B R, Bickford A A, et al. Hexamita meleagridis (Spironucleus meleagridis) infection in chukar partridges associated with high mortality and intracellular trophozoites[J]. Avian diseases, 2004, 48(3): 706-710.